

Cysteine dioxygenase is essential for mouse sperm osmoadaptation and male fertility
- PMID: 29604178
- PMCID: PMC5992081
- DOI: 10.1111/febs.14449
Cysteine dioxygenase is essential for mouse sperm osmoadaptation and male fertility
Abstract
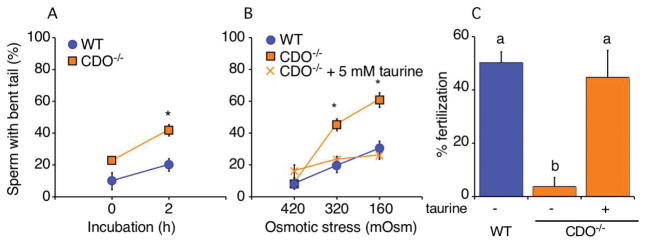
Sperm entering the epididymis are immotile and cannot respond to stimuli that will enable them to fertilize. The epididymis is a highly complex organ, with multiple histological zones and cell types that together change the composition and functional abilities of sperm through poorly understood mechanisms. Sperm take up taurine during epididymal transit, which may play antioxidant or osmoregulatory roles. Cysteine dioxygenase (CDO) is a critical enzyme for taurine synthesis. A previous study reported that male CDO-/- mice exhibit idiopathic infertility, prompting us to investigate the functions of CDO in male fertility. Immunoblotting and quantitative reverse transcription-polymerase chain reaction analysis of epididymal segments showed that androgen-dependent CDO expression was highest in the caput epididymidis. CDO-/- mouse sperm demonstrated a severe lack of in vitro fertilization ability. Acrosome exocytosis and tyrosine phosphorylation profiles in response to stimuli were normal, suggesting normal functioning of pathways associated with capacitation. CDO-/- sperm had a slight increase in head abnormalities. Taurine and hypotaurine concentrations in CDO-/- sperm decreased in the epididymal intraluminal fluid and sperm cytosol. We found no evidence of antioxidant protection against lipid peroxidation. However, CDO-/- sperm exhibited severe defects in volume regulation, swelling in response to the relatively hypo-osmotic conditions found in the female reproductive tract. Our findings suggest that epididymal CDO plays a key role in post-testicular sperm maturation, enabling sperm to osmoregulate as they transition from the male to the female reproductive tract, and provide new understanding of the compartmentalized functions of the epididymis.
Keywords: epididymal maturation; fertilization; sperm; taurine.
© 2018 Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare no conflicts of interest associated with this manuscript.
Figures

Similar articles
-
Epididymal cysteine-rich secretory proteins are required for epididymal sperm maturation and optimal sperm function.Mol Hum Reprod. 2018 Mar 1;24(3):111-122. doi: 10.1093/molehr/gay001. Mol Hum Reprod. 2018. PMID: 29361143
-
Junctional adhesion molecule A: expression in the murine epididymal tract and accessory organs and acquisition by maturing sperm.Mol Hum Reprod. 2017 Feb 10;23(2):132-140. doi: 10.1093/molehr/gaw082. Mol Hum Reprod. 2017. PMID: 28062807 Free PMC article.
-
Caput Ligation Renders Immature Mouse Sperm Motile and Capable to Undergo cAMP-Dependent Phosphorylation.Int J Mol Sci. 2021 Sep 23;22(19):10241. doi: 10.3390/ijms221910241. Int J Mol Sci. 2021. PMID: 34638585 Free PMC article.
-
Relevance of CRISP proteins for epididymal physiology, fertilization, and fertility.Andrology. 2019 Sep;7(5):610-617. doi: 10.1111/andr.12638. Epub 2019 Jun 19. Andrology. 2019. PMID: 31218833 Review.
-
Understanding human thiol dioxygenase enzymes: structure to function, and biology to pathology.Int J Exp Pathol. 2017 Apr;98(2):52-66. doi: 10.1111/iep.12222. Epub 2017 Apr 24. Int J Exp Pathol. 2017. PMID: 28439920 Free PMC article. Review.
Cited by
-
Cloning, bioinformatics analysis and expression of the cysteine dioxygenase type 1 (CDO1) gene in domestic yak.Front Vet Sci. 2024 Oct 18;11:1488782. doi: 10.3389/fvets.2024.1488782. eCollection 2024. Front Vet Sci. 2024. PMID: 39493813 Free PMC article.
-
The role of taurine in male reproduction: Physiology, pathology and toxicology.Front Endocrinol (Lausanne). 2023 Jan 18;14:1017886. doi: 10.3389/fendo.2023.1017886. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 36742382 Free PMC article. Review.
-
MiRNA-34c Regulates Bovine Sertoli Cell Proliferation, Gene Expression, and Apoptosis by Targeting the AXL Gene.Animals (Basel). 2021 Aug 13;11(8):2393. doi: 10.3390/ani11082393. Animals (Basel). 2021. PMID: 34438849 Free PMC article.
-
Monitoring of taurine dietary supplementation effect on parameters of Duroc boar ejaculate in summer season.PLoS One. 2024 Jan 25;19(1):e0288317. doi: 10.1371/journal.pone.0288317. eCollection 2024. PLoS One. 2024. PMID: 38271350 Free PMC article.
-
A proposed role for CDO1 in CNS development: Three children with rare missense variants and a neurological phenotype.HGG Adv. 2025 Apr 10;6(2):100417. doi: 10.1016/j.xhgg.2025.100417. Epub 2025 Feb 13. HGG Adv. 2025. PMID: 39949058 Free PMC article.
References
-
- Cooper TG, Yeung CH. Acquisition of volume regulatory response of sperm upon maturation in the epididymis and the role of the cytoplasmic droplet. Microsc Res Tech. 2003;61:28–38. - PubMed
-
- Yeung CH, Wagenfeld A, Nieschlag E, Cooper TG. The cause of infertility of male c-ros tyrosine kinase receptor knockout mice. Biol Reprod. 2000;63:612–8. - PubMed
-
- Yeung CH, Sonnenberg-Riethmacher E, Cooper TG. Infertile spermatozoa of c-ros tyrosine kinase receptor knockout mice show flagellar angulation and maturational defects in cell volume regulatory mechanisms. Biol Reprod. 1999;61:1062–9. - PubMed
-
- Yeung CH, Anapolski M, Sipila P, Wagenfeld A, Poutanen M, Huhtaniemi I, Nieschlag E, Cooper TG. Sperm volume regulation: maturational changes in fertile and infertile transgenic mice and association with kinematics and tail angulation. Biol Reprod. 2002;67:269–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
