

Translation-relevant EEG phenotypes in a mouse model of Fragile X Syndrome
- PMID: 29605426
- PMCID: PMC5969806
- DOI: 10.1016/j.nbd.2018.03.012
Translation-relevant EEG phenotypes in a mouse model of Fragile X Syndrome
Abstract
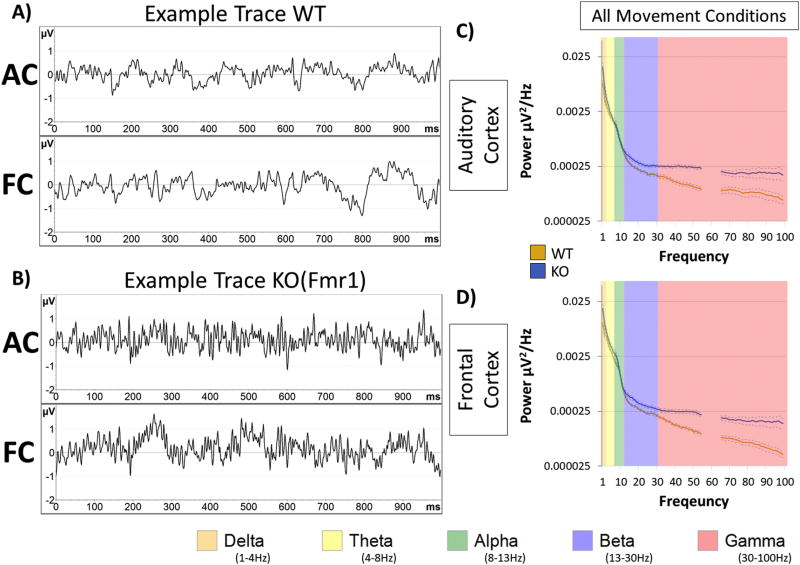
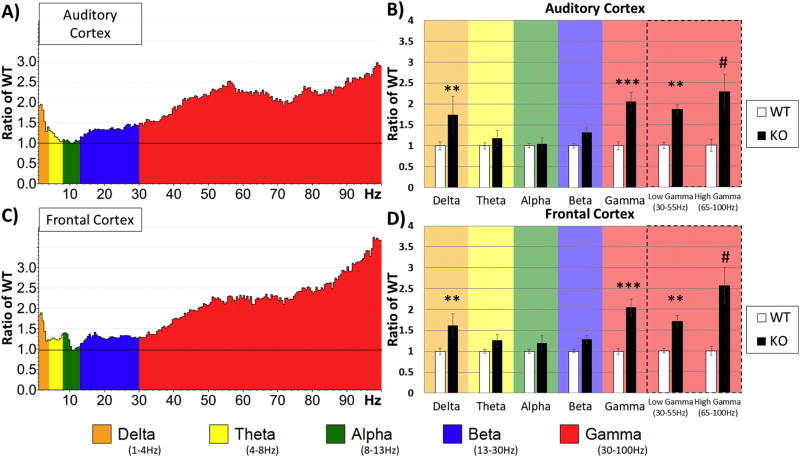
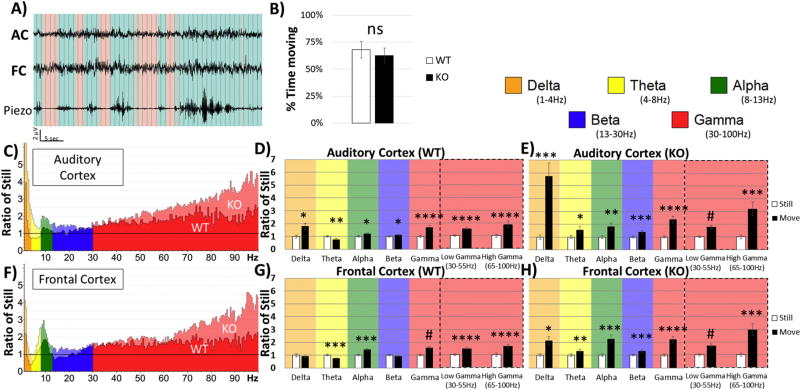
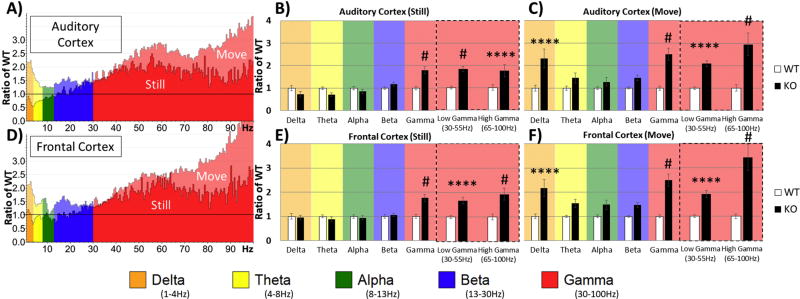
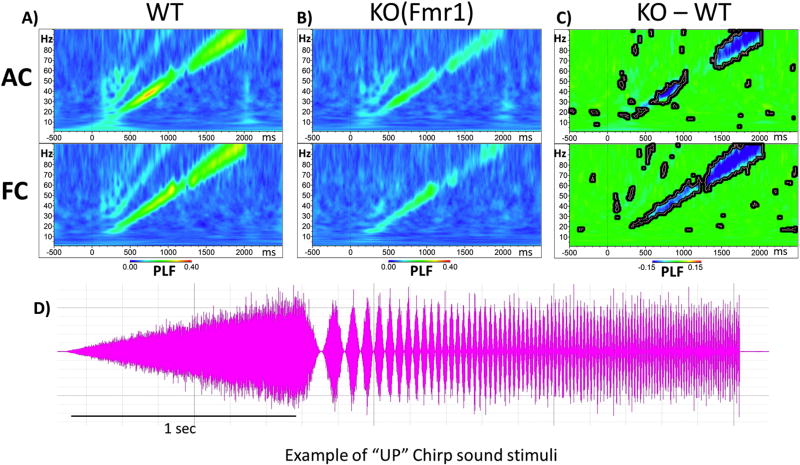
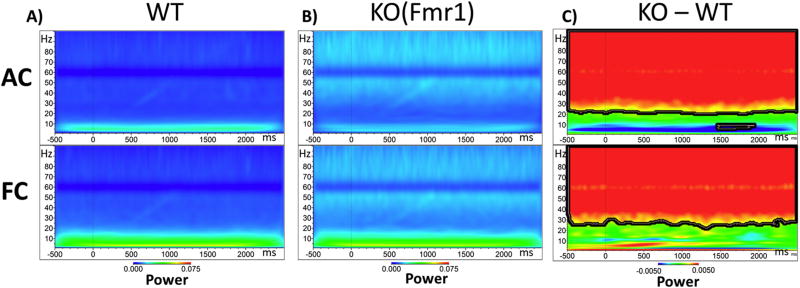
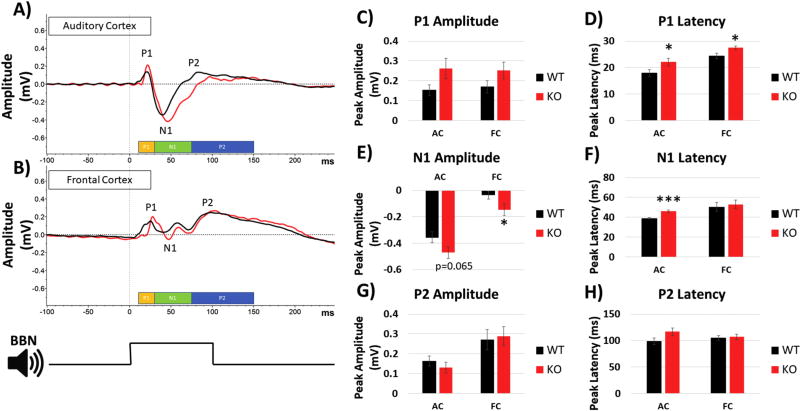
Identification of comparable biomarkers in humans and validated animal models will facilitate pre-clinical to clinical therapeutic pipelines to treat neurodevelopmental disorders. Fragile X Syndrome (FXS) is a leading known genetic cause of intellectual disability with symptoms that include increased anxiety, social and sensory processing deficits. Recent EEG studies in humans with FXS have identified neural oscillation deficits that include enhanced resting state gamma power and reduced inter-trial coherence of sound evoked gamma oscillations. To determine if analogous phenotypes are present in an animal model of FXS, we recorded EEGs in awake, freely moving Fmr1 knock out (KO) mice using similar stimuli as in the human studies. We report remarkably similar neural oscillation phenotypes in the Fmr1 KO mouse including enhanced resting state gamma power and reduced evoked gamma synchronization. The gamma band inter-trial coherence of neural response was reduced in both auditory and frontal cortex of Fmr1 KO mice stimulated with a sound whose envelope was modulated from 1 to 100 Hz, similar to that seen in humans with FXS. These deficits suggest a form of enhanced 'resting state noise' that interferes with the ability of the circuit to mount a synchronized response to sensory input, predicting specific sensory and cognitive deficits in FXS. The abnormal gamma oscillations are consistent with parvalbumin neuron and perineuronal net deficits seen in the Fmr1 KO mouse auditory cortex indicating that the EEG biomarkers are not only clinically relevant, but could also be used to probe cellular and circuit mechanisms of sensory hypersensitivity in FXS.
Keywords: Auditory cortex; Autism; EEG; Fragile X Syndrome; Frontal cortex; Neural oscillations; Sensory hypersensitivity.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
References
-
- Abbeduto L, Hagerman RJ. Language and communication in fragile X syndrome. Developmental Disabilities Research Reviews. 1997;3(4):313–322.
-
- Bakker CE, Verheij C, Willemsen R, Vanderhelm R, Oerlemans F, Vermey M, Bygrave A, Hoogeveen AT, Oostra BA, Reyniers E, Deboulle K, Dhooge R, Cras P, Vanvelzen D, Nagels G, Martin JJ, Dedeyn PP, Darby JK, Willems PJ. Fmr1 knockout mice - a model to study fragile-x mental-retardation. Cell. 1994;78:23–33. - PubMed
-
- Berry-Kravis E. Epilepsy in fragile X syndrome. Dev Med Child Neurol. 2002;44:724–728. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
