

FINO2 initiates ferroptosis through GPX4 inactivation and iron oxidation
- PMID: 29610484
- PMCID: PMC5899674
- DOI: 10.1038/s41589-018-0031-6
FINO2 initiates ferroptosis through GPX4 inactivation and iron oxidation
Abstract
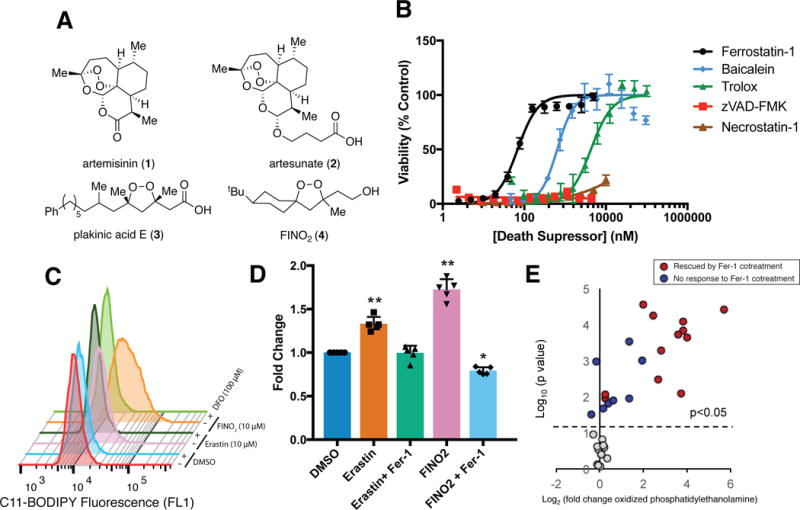
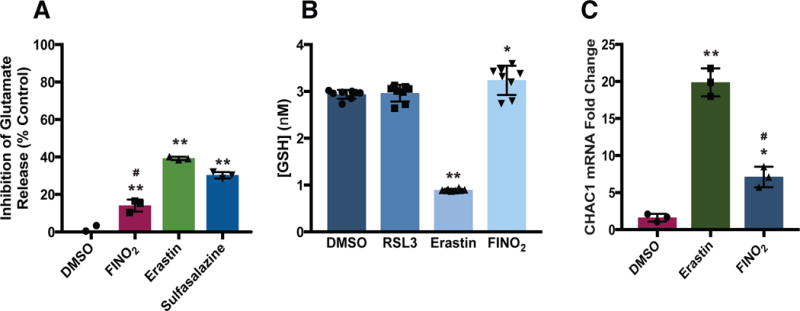
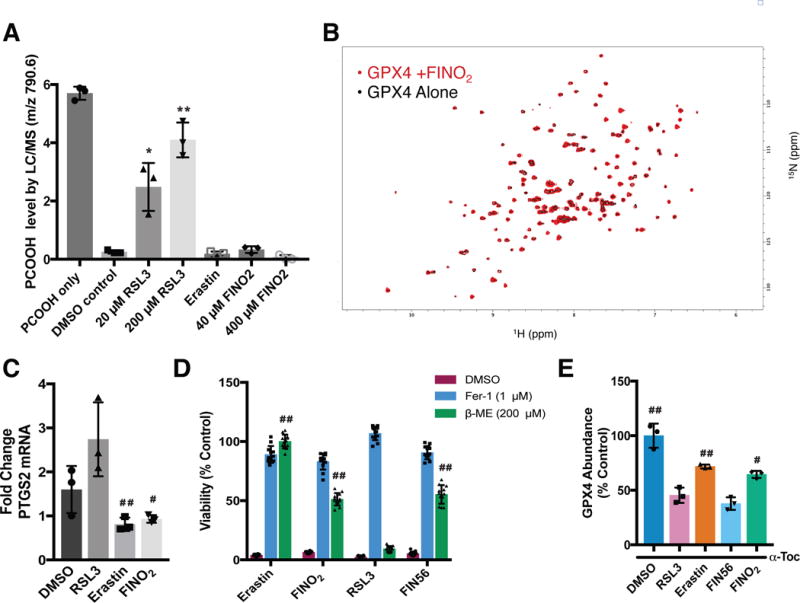
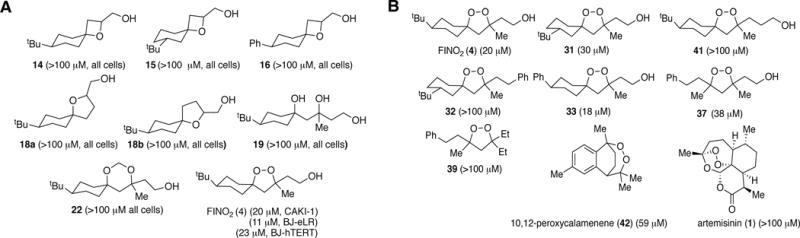
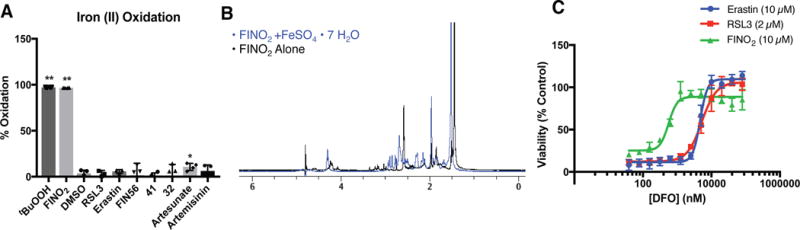
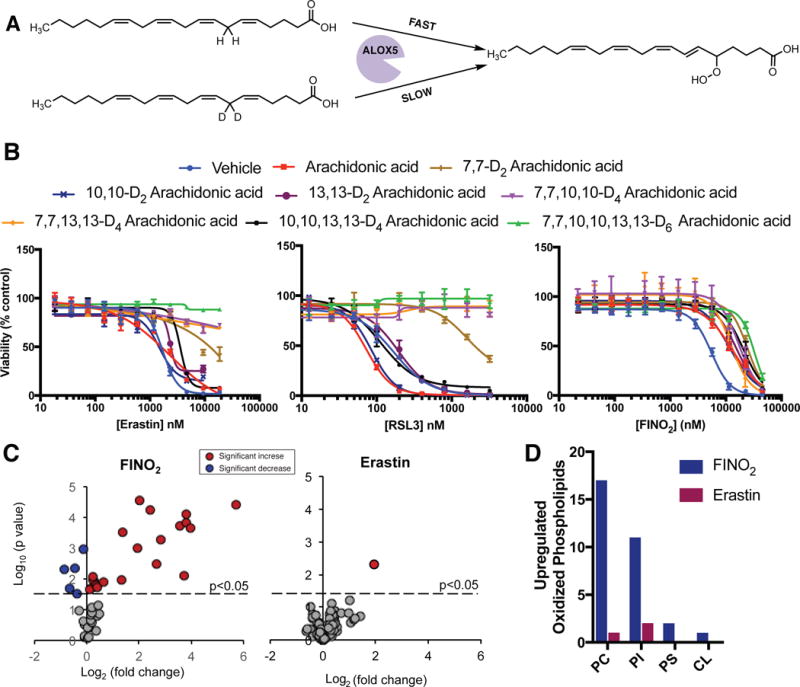
Ferroptosis is a non-apoptotic form of regulated cell death caused by the failure of the glutathione-dependent lipid-peroxide-scavenging network. FINO2 is an endoperoxide-containing 1,2-dioxolane that can initiate ferroptosis selectively in engineered cancer cells. We investigated the mechanism and structural features necessary for ferroptosis initiation by FINO2. We found that FINO2 requires both an endoperoxide moiety and a nearby hydroxyl head group to initiate ferroptosis. In contrast to previously described ferroptosis inducers, FINO2 does not inhibit system xc- or directly target the reducing enzyme GPX4, as do erastin and RSL3, respectively, nor does it deplete GPX4 protein, as does FIN56. Instead, FINO2 both indirectly inhibits GPX4 enzymatic function and directly oxidizes iron, ultimately causing widespread lipid peroxidation. These findings suggest that endoperoxides such as FINO2 can initiate a multipronged mechanism of ferroptosis.
Conflict of interest statement
M.S. Shchepinov declares a competing financial interest as the Chief Scientific Officer of Retrotrope, Inc., all other authors declare no competing financial interests.
Figures
References
-
- Lockshin RA, Zakeri Z. Programmed cell death and apoptosis: origins of the theory. Nat Rev Mol Cell Biol. 2001;2:545–550. - PubMed
Methods-only References
-
- Folch J, Lees M, Sloane Stanley GH. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem. 1957;226:497–509. - PubMed
-
- Boettcher C, Pries C, Vangent CM. A rapid and sensitive sub-micro phosphorus determination. Anal Chim Acta. 1961;24:203–204.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
