

Impact of F1Fo-ATP-synthase dimer assembly factors on mitochondrial function and organismic aging
- PMID: 29610761
- PMCID: PMC5878687
- DOI: 10.15698/mic2018.04.625
Impact of F1Fo-ATP-synthase dimer assembly factors on mitochondrial function and organismic aging
Abstract
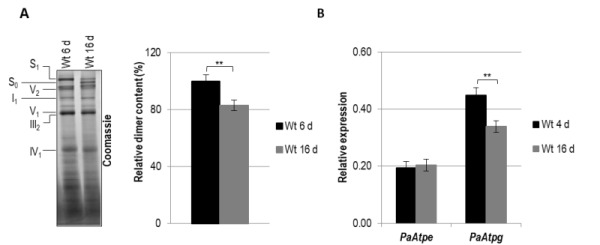
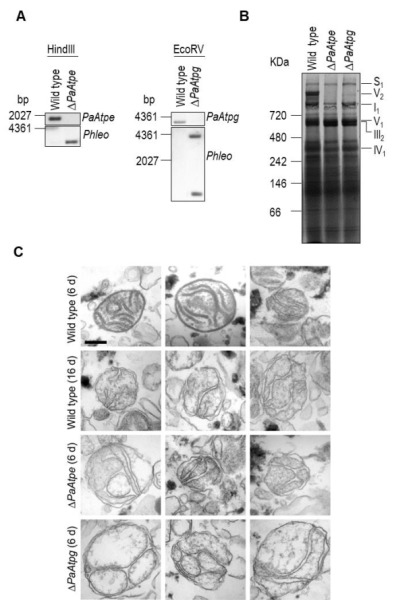
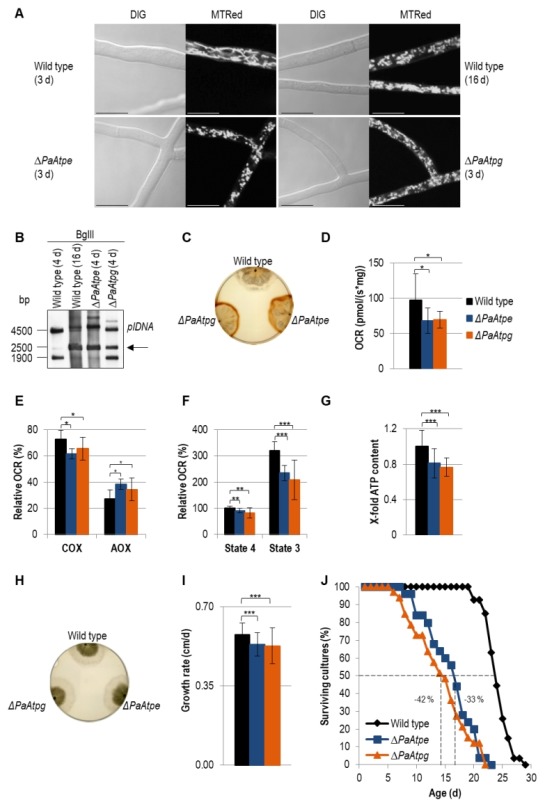
In aerobic organisms, mitochondrial F1Fo-ATP-synthase is the major site of ATP production. Beside this fundamental role, the protein complex is involved in shaping and maintenance of cristae. Previous electron microscopic studies identified the dissociation of F1Fo-ATP-synthase dimers and oligomers during organismic aging correlating with a massive remodeling of the mitochondrial inner membrane. Here we report results aimed to experimentally proof this impact and to obtain further insights into the control of these processes. We focused on the role of the two dimer assembly factors PaATPE and PaATPG of the aging model Podospora anserina. Ablation of either protein strongly affects mitochondrial function and leads to an accumulation of senescence markers demonstrating that the inhibition of dimer formation negatively influences vital functions and accelerates organismic aging. Our data validate a model that links mitochondrial membrane remodeling to aging and identify specific molecular components triggering this process.
Keywords: F1Fo-ATP-synthase; aging; membranes; mitochondria; remodeling.
Conflict of interest statement
Conflict of interest: The authors declare that they have no conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources