

Pseudomonas aeruginosa and Klebsiella pneumoniae Adaptation to Innate Immune Clearance Mechanisms in the Lung
- PMID: 29617698
- PMCID: PMC6785651
- DOI: 10.1159/000487515
Pseudomonas aeruginosa and Klebsiella pneumoniae Adaptation to Innate Immune Clearance Mechanisms in the Lung
Abstract
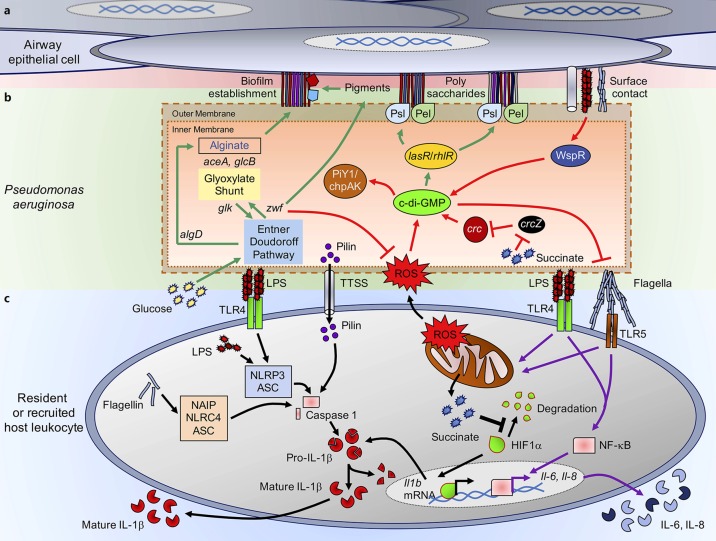
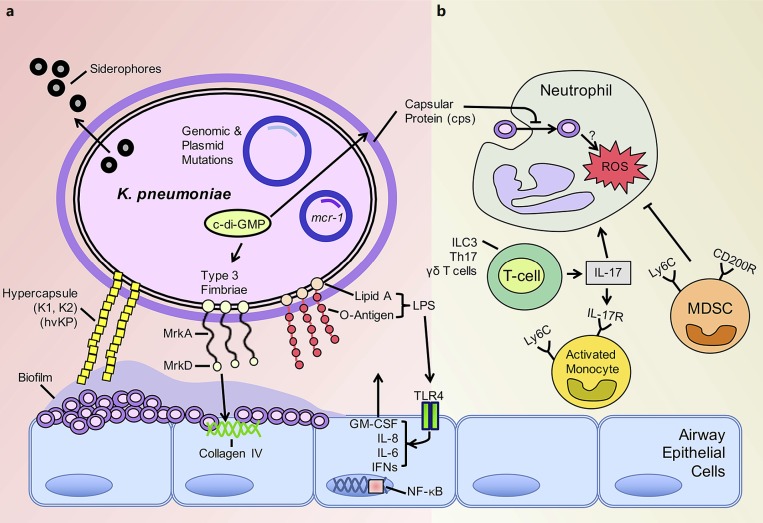
Many different species of gram-negative bacteria are associated with infection in the lung, causing exacerbations of chronic obstructive pulmonary disease, cystic fibrosis (CF), and ventilator-associated pneumonias. These airway pathogens must adapt to common host clearance mechanisms that include killing by antimicrobial peptides, antibiotics, oxidative stress, and phagocytosis by leukocytes. Bacterial adaptation to the host is often evident phenotypically, with increased extracellular polysaccharide production characteristic of some biofilm-associated organisms. Given the relatively limited repertoire of bacterial strategies to elude airway defenses, it seems likely that organisms sharing the same ecological niche might also share common strategies to persistently infect the lung. In this review, we will highlight some of the major factors responsible for the adaptation of Pseudomonas aeruginosa to the lung, addressing how growth in biofilms enables persistent infection, relevant to, but not limited to, the pathogenesis of infection in CF. In contrast, we will discuss how carbapenem-resistant Klebsiella pneumoniae evade immune clearance, an organism often associated with ventilator-associated pneumonia and health-care-acquired pneumonias, but not a typical pathogen in CF.
Keywords: Bacterial adaptation; Bacterial infection; Biofilm; Cystic fibrosis; Immune evasion; Inflammasome; Klebsiella pneumoniae; Pathogen-associated molecular patterns; Pseudomonas aeruginosa; Reactive oxygen species.
© 2018 S. Karger AG, Basel.
Figures
References
-
- Elborn JS. Cystic fibrosis. Lancet. 2016;388:2519–2531. - PubMed
-
- Gutierrez HH, Sanchez I, Schidlow DV. Cystic fibrosis care in Chile. Curr Opin Pulm Med. 2009;15:632–637. - PubMed
-
- Silva Filho LV, Castanos C, Ruiz HH. Cystic fibrosis in Latin America - improving the awareness. J Cyst Fibros. 2016;15:791–793. - PubMed
-
- Masekela R, Zampoli M, Westwood AT, White DA, Green RJ, Olorunju S, Kwofie-Mensah M. Phenotypic expression of the 3120 + 1G>A mutation in non-Caucasian children with cystic fibrosis in South Africa. J Cyst Fibros. 2013;12:363–366. - PubMed
-
- Westwood T, Henderson B, Ramsay M, Medical and Scientific Advisory Committee of the South African Cystic Fibrosis Association Diagnosing cystic fibrosis in South Africa. S Afr Med J. 2006;96:304–306. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
