

NADPH oxidase activation regulates apoptotic neutrophil clearance by murine macrophages
- PMID: 29618478
- PMCID: PMC5969376
- DOI: 10.1182/blood-2017-09-809004
NADPH oxidase activation regulates apoptotic neutrophil clearance by murine macrophages
Abstract
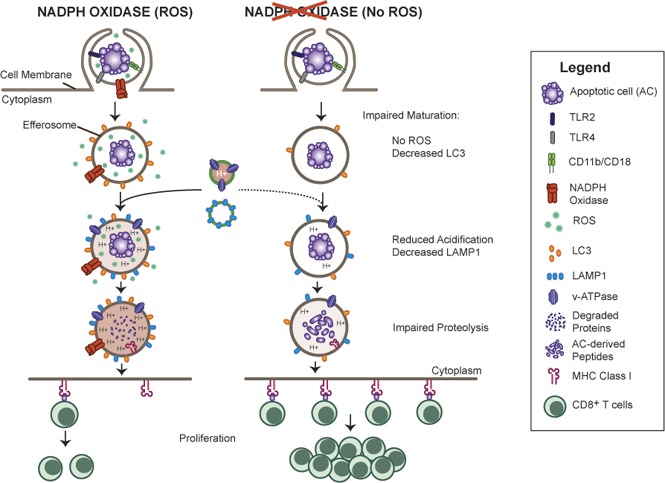
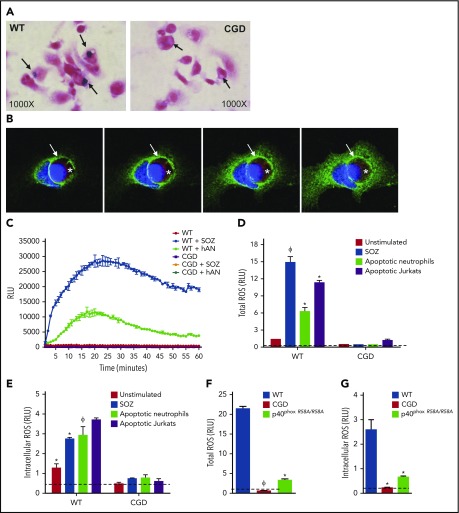
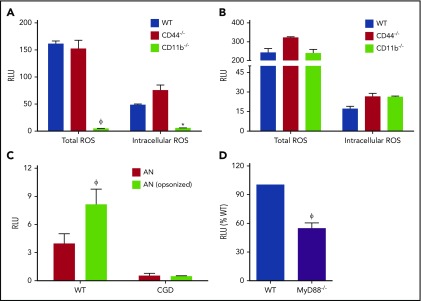
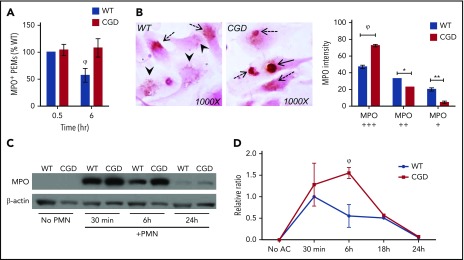
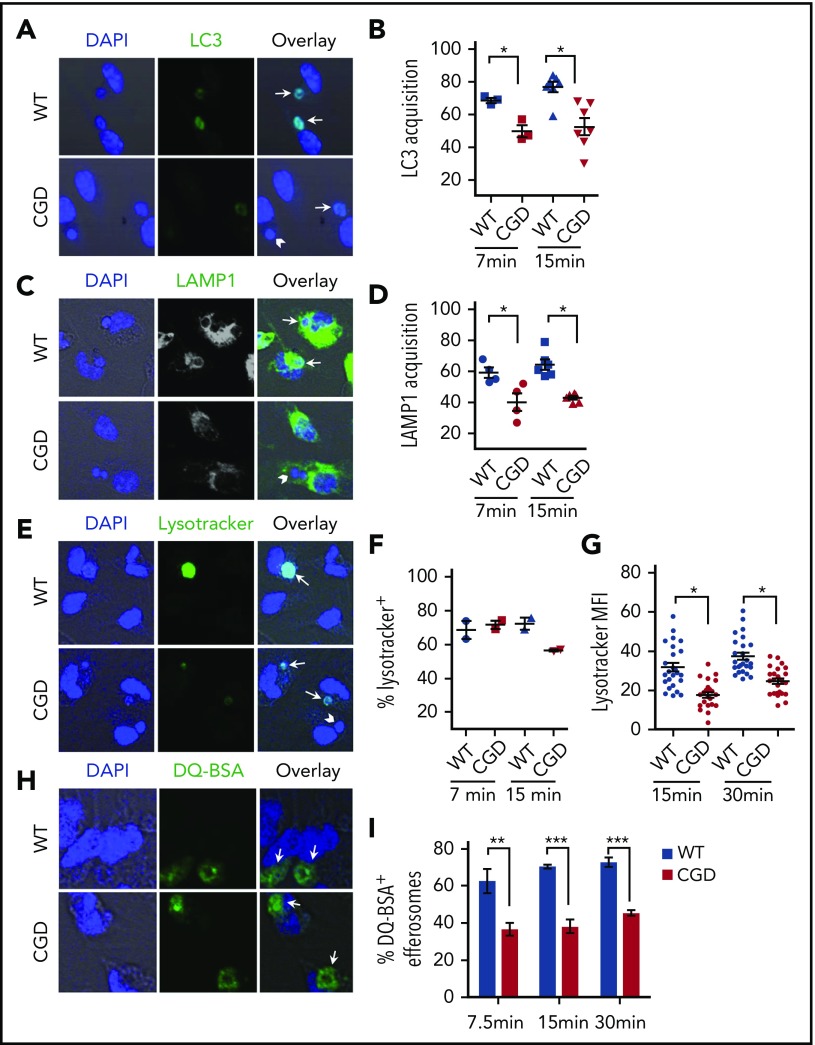
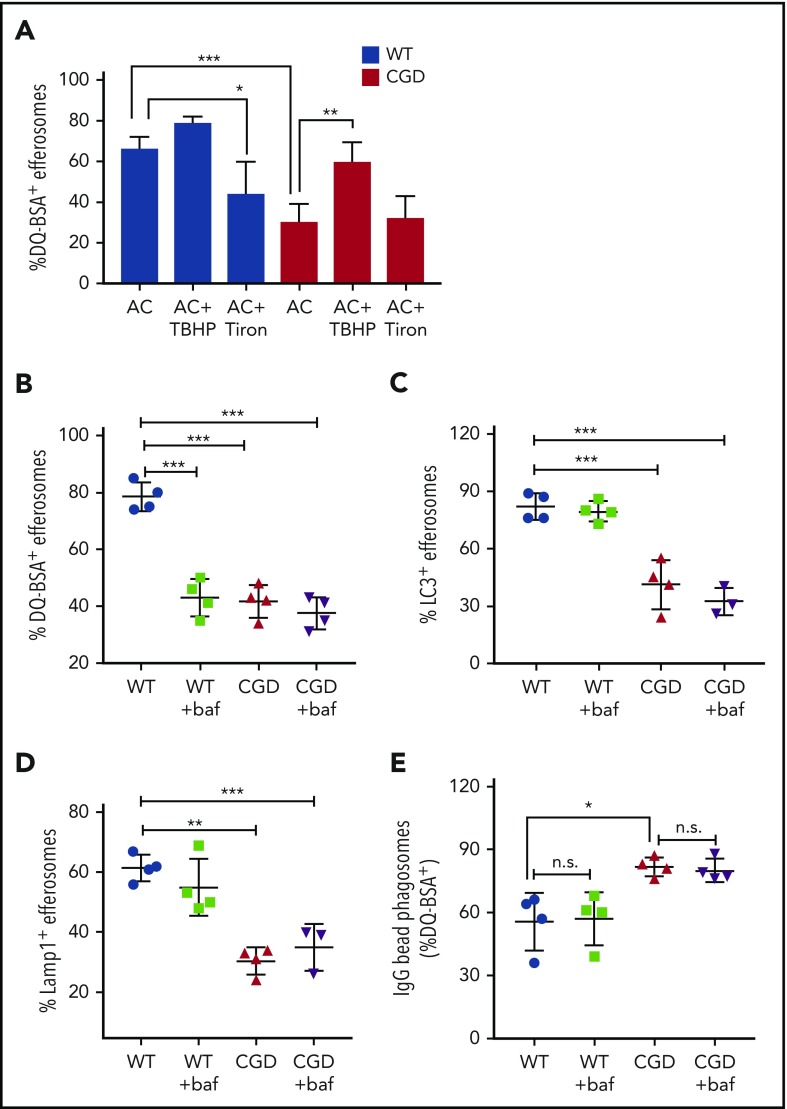
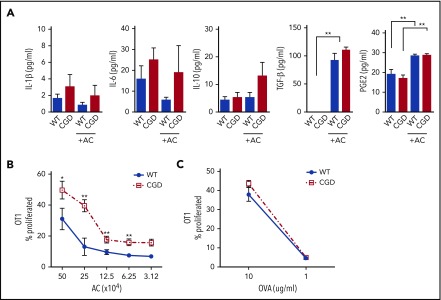
The phagocyte reduced NAD phosphate (NADPH) oxidase generates superoxide, the precursor to reactive oxygen species (ROS) that has both antimicrobial and immunoregulatory functions. Inactivating mutations in NADPH oxidase alleles cause chronic granulomatous disease (CGD), characterized by enhanced susceptibility to life-threatening microbial infections and inflammatory disorders; hypomorphic NADPH oxidase alleles are associated with autoimmunity. Impaired apoptotic cell (AC) clearance is implicated as an important contributing factor in chronic inflammation and autoimmunity, but the role of NADPH oxidase-derived ROS in this process is incompletely understood. Here, we demonstrate that phagocytosis of AC (efferocytosis) potently activated NADPH oxidase in mouse peritoneal exudate macrophages (PEMs). ROS generation was dependent on macrophage CD11b, Toll-like receptor 2 (TLR2), TLR4, and myeloid differentiation primary response 88 (MyD88), and was also regulated by phosphatidylinositol 3-phosphate binding to the p40 phox oxidase subunit. Maturation of efferosomes containing apoptotic neutrophils was significantly delayed in CGD PEMs, including acidification and acquisition of proteolytic activity, and was associated with slower digestion of apoptotic neutrophil proteins. Treatment of wild-type macrophages with the vacuolar-type H+ ATPase inhibitor bafilomycin also delayed proteolysis within efferosomes, showing that luminal acidification was essential for efficient digestion of efferosome proteins. Finally, cross-presentation of AC-associated antigens by CGD PEMs to CD8 T cells was increased. These studies unravel a key role for the NADPH oxidase in the disposal of ACs by inflammatory macrophages. The oxidants generated promote efferosome maturation and acidification that facilitate the degradation of ingested ACs.
© 2018 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
References
-
- Holmdahl R, Sareila O, Olsson LM, Bäckdahl L, Wing K. Ncf1 polymorphism reveals oxidative regulation of autoimmune chronic inflammation. Immunol Rev. 2016;269(1):228-247. - PubMed
-
- Lam GY, Huang J, Brumell JH. The many roles of NOX2 NADPH oxidase-derived ROS in immunity. Semin Immunopathol. 2010;32(4):415-430. - PubMed
-
- Dinauer M, Newburger P, Borregaard N. The phagocyte system and disorders of granulopoiesis and granulocyte function. In: Nathan DG, Orkin SH, eds. Nathan and Oski’s hematology of infancy and childhood. Philadelphia, PA: Elsevier/Saunders; 2014.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
