

Biology in Bloom: A Primer on the Arabidopsis thaliana Model System
- PMID: 29618591
- PMCID: PMC5887134
- DOI: 10.1534/genetics.118.300755
Biology in Bloom: A Primer on the Arabidopsis thaliana Model System
Abstract
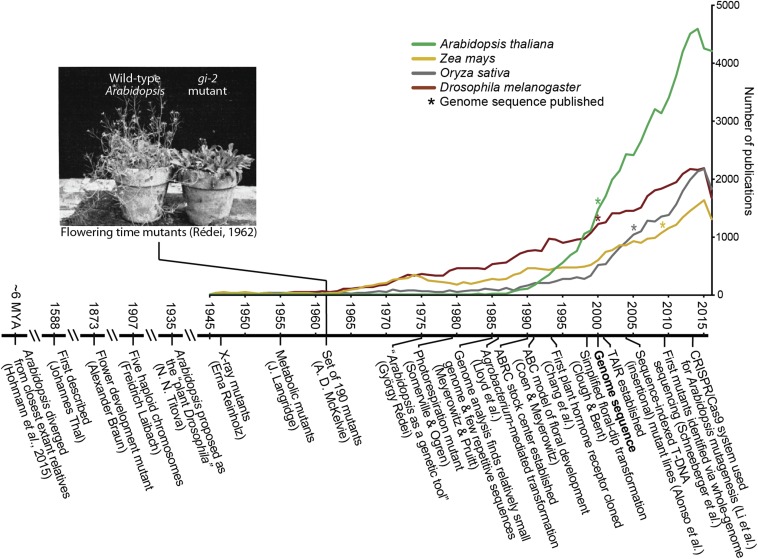
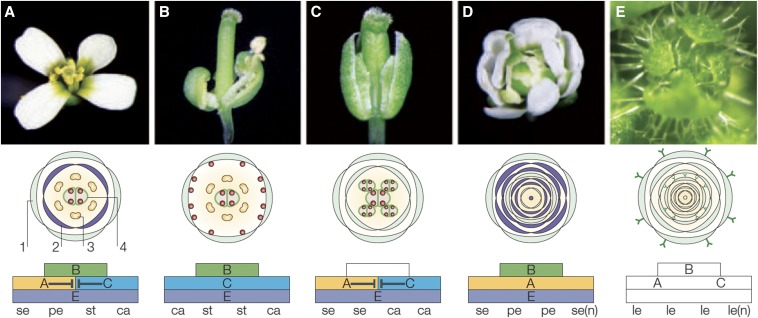
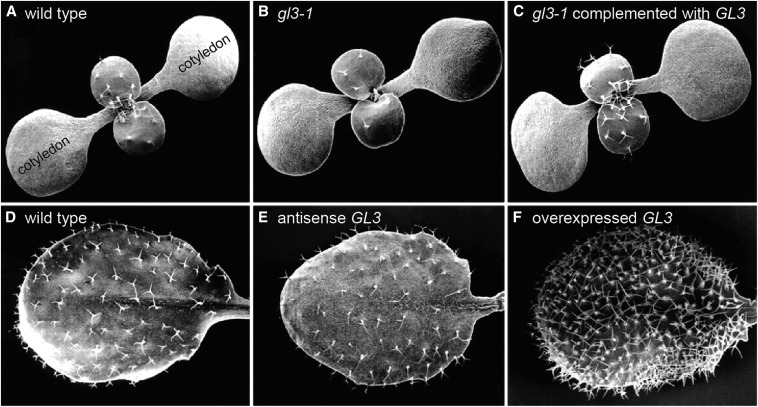
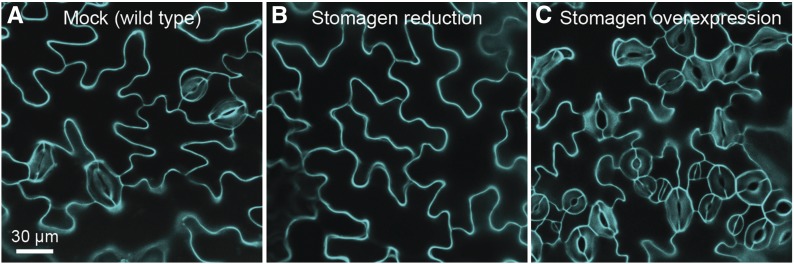
Arabidopsis thaliana could have easily escaped human scrutiny. Instead, Arabidopsis has become the most widely studied plant in modern biology despite its absence from the dinner table. Pairing diminutive stature and genome with prodigious resources and tools, Arabidopsis offers a window into the molecular, cellular, and developmental mechanisms underlying life as a multicellular photoautotroph. Many basic discoveries made using this plant have spawned new research areas, even beyond the verdant fields of plant biology. With a suite of resources and tools unmatched among plants and rivaling other model systems, Arabidopsis research continues to offer novel insights and deepen our understanding of fundamental biological processes.
Keywords: model organism; reference plant.
Copyright © 2018 by the Genetics Society of America.
Figures



References
-
- Allen E., Xie Z., Gustafson A. M., Carrington J. C., 2005. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121: 207–221. - PubMed
-
- Alonso J. M., Stepanova A. N., Leisse T. J., Kim C. J., Chen H., et al. , 2003. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657. - PubMed
-
- Arabidopsis Genome Initiative , 2000. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408: 796–815. - PubMed
-
- Baerenfaller K., Grossmann J., Grobei M. A., Hull R., Hirsch-Hoffmann M., et al. , 2008. Genome-scale proteomics reveals Arabidopsis thaliana gene models and proteome dynamics. Science 320: 938–941. - PubMed
-
- Bauwe H., Hagemann M., Fernie A. R., 2010. Photorespiration: players, partners and origin. Trends Plant Sci. 15: 330–336. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
