

The inflammasome potentiates influenza/Staphylococcus aureus superinfection in mice
- PMID: 29618653
- PMCID: PMC5928863
- DOI: 10.1172/jci.insight.97470
The inflammasome potentiates influenza/Staphylococcus aureus superinfection in mice
Abstract
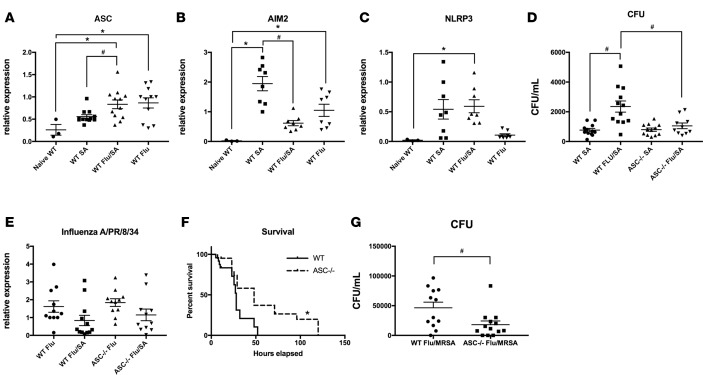
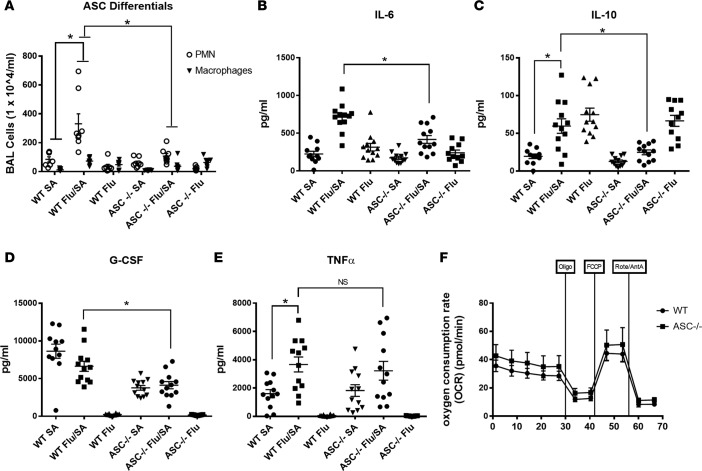
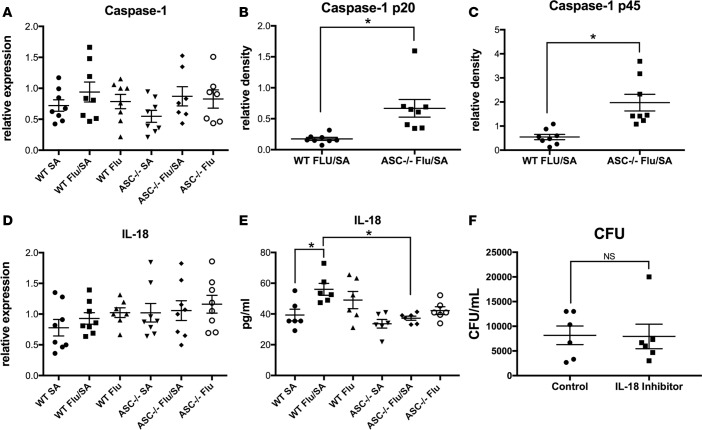
Secondary bacterial respiratory infections are commonly associated with both acute and chronic lung injury. Influenza complicated by bacterial pneumonia is an effective model to study host defense during pulmonary superinfection due to its clinical relevance. Multiprotein inflammasomes are responsible for IL-1β production in response to infection and drive tissue inflammation. In this study, we examined the role of the inflammasome during viral/bacterial superinfection. We demonstrate that ASC-/- mice are protected from bacterial superinfection and produce sufficient quantities of IL-1β through an apoptosis-associated speck-like protein containing CARD (ASC) inflammasome-independent mechanism. Despite the production of IL-1β by ASC-/- mice in response to bacterial superinfection, these mice display decreased lung inflammation. A neutrophil elastase inhibitor blocked ASC inflammasome-independent production of IL-1β and the IL-1 receptor antagonist, anakinra, confirmed that IL-1 remains crucial to the clearance of bacteria during superinfection. Delayed inhibition of NLRP3 during influenza infection by MCC950 decreases bacterial burden during superinfection and leads to decreased inflammatory cytokine production. Collectively, our results demonstrate that ASC augments the clearance of bacteria, but can also contribute to inflammation and mortality. ASC should be considered as a therapeutic target to decrease morbidity and mortality during bacterial superinfection.
Keywords: Bacterial infections; Immunology; Influenza; Pulmonology.
Conflict of interest statement
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
