

Distributed hepatocytes expressing telomerase repopulate the liver in homeostasis and injury
- PMID: 29618815
- PMCID: PMC5895494
- DOI: 10.1038/s41586-018-0004-7
Distributed hepatocytes expressing telomerase repopulate the liver in homeostasis and injury
Abstract
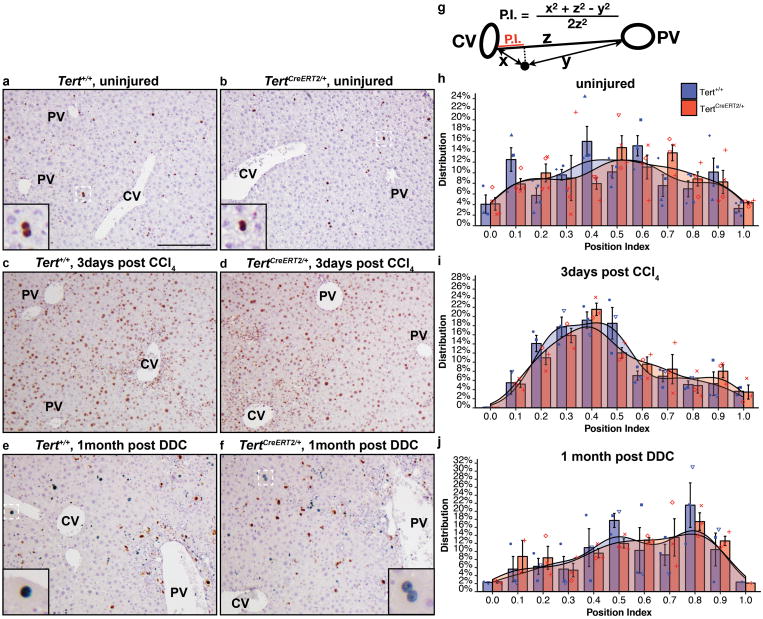
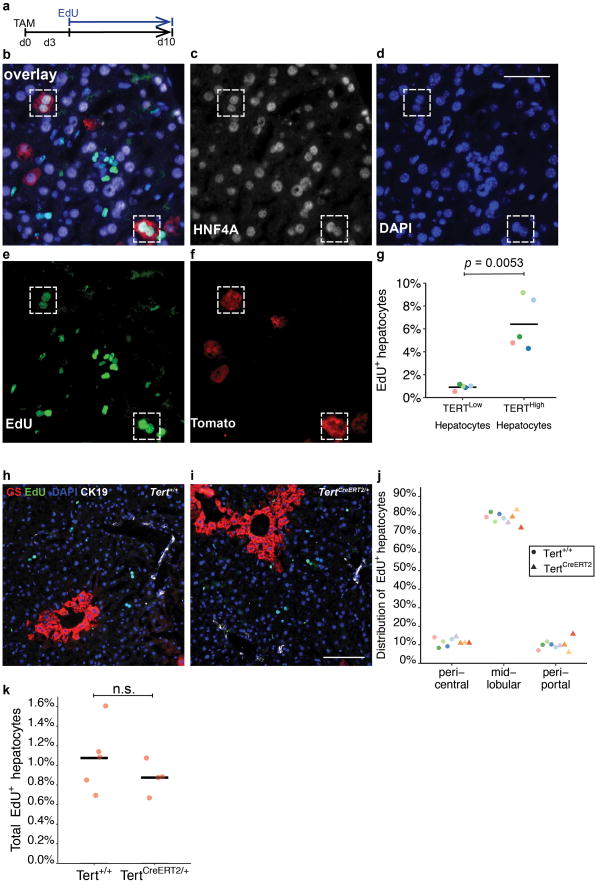
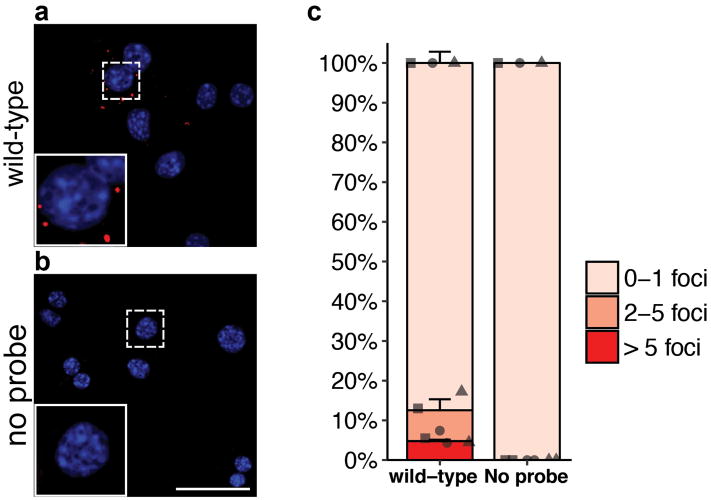
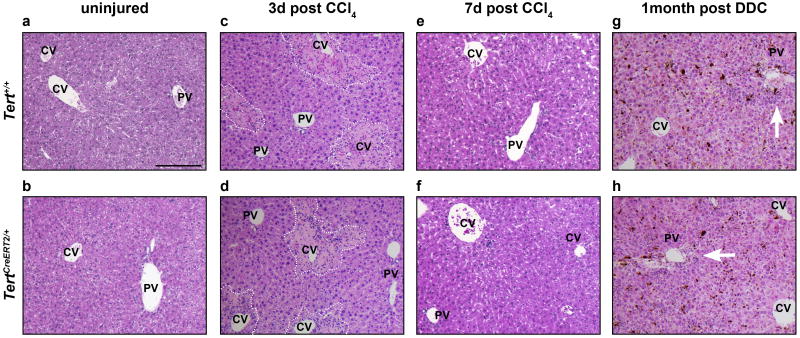
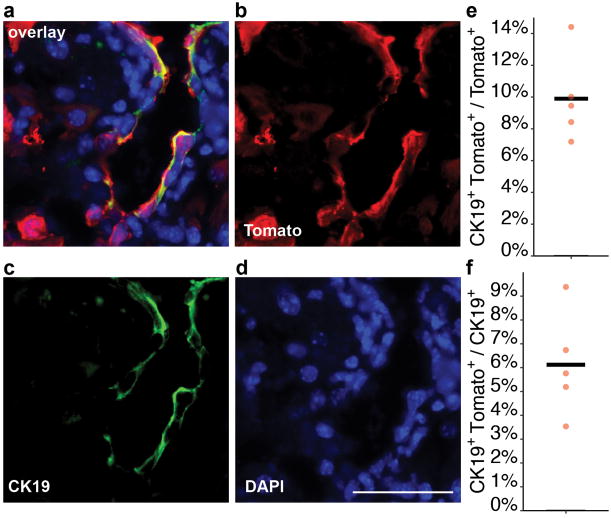
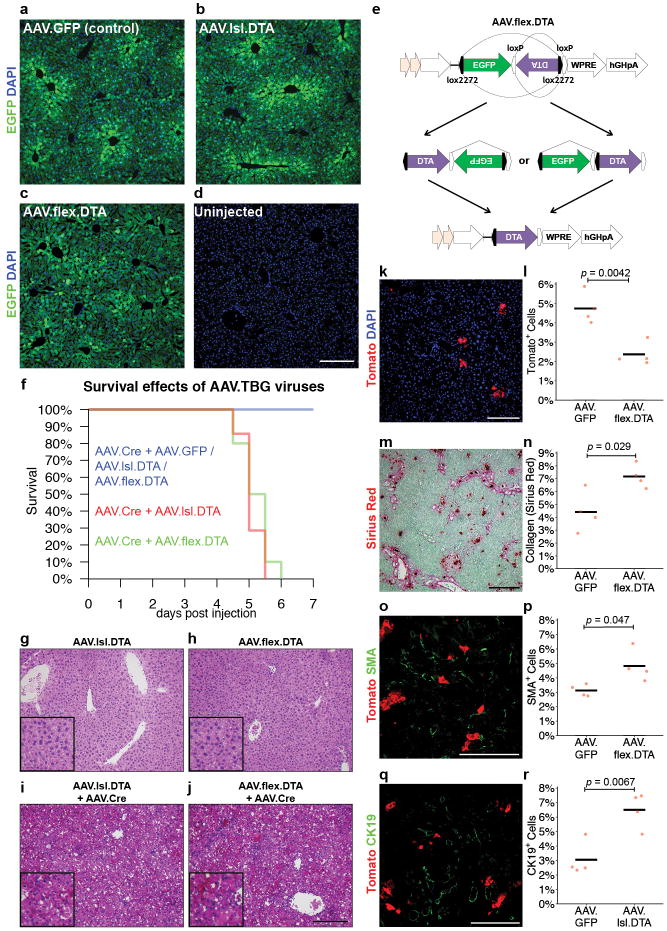
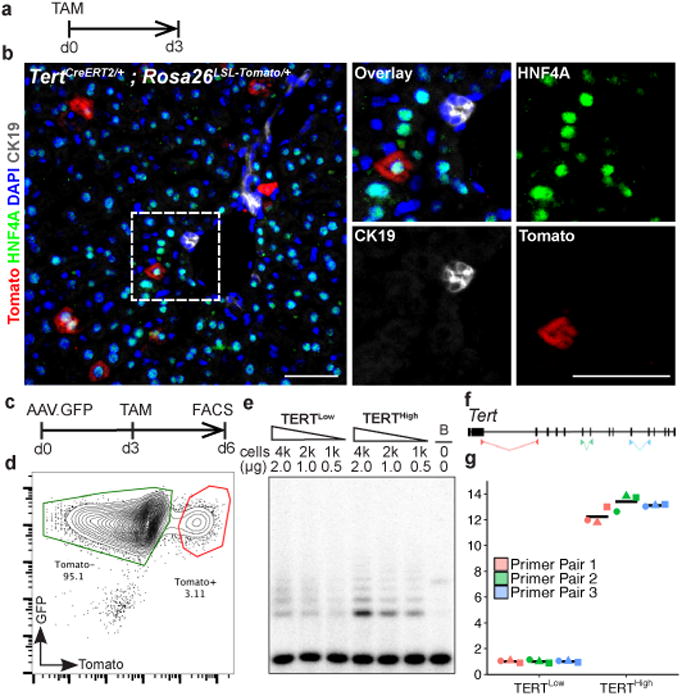
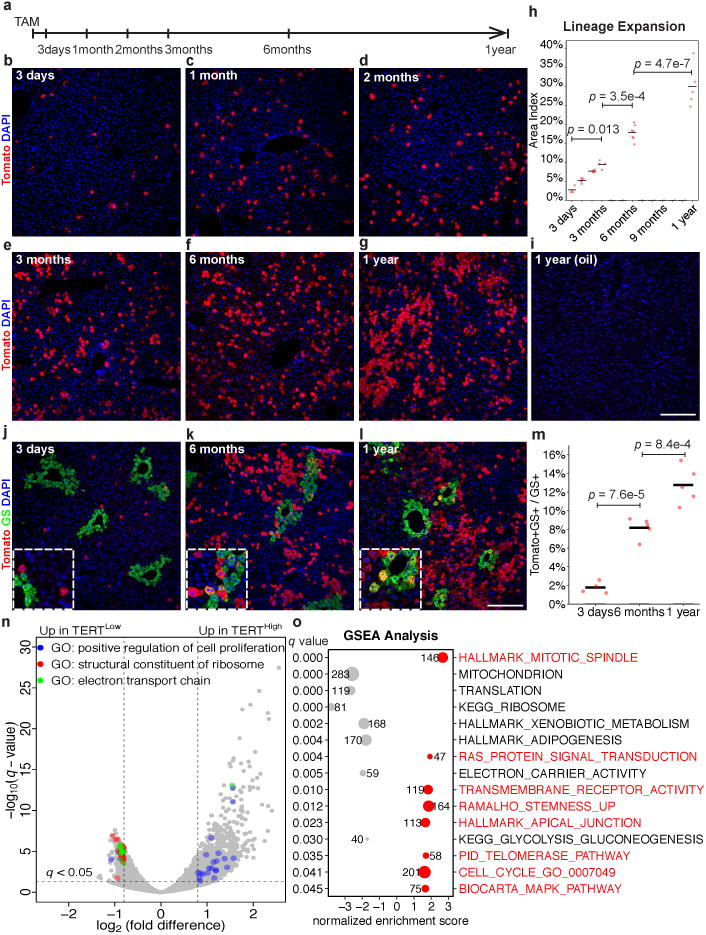
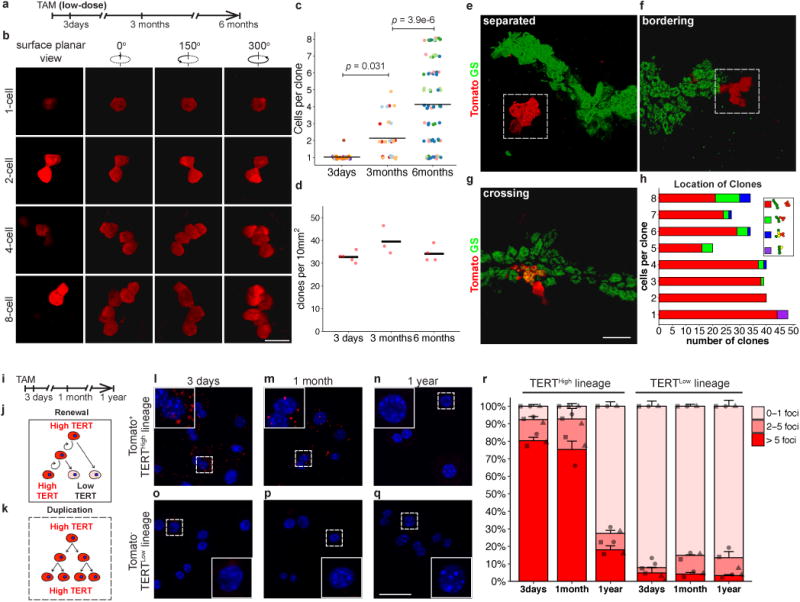
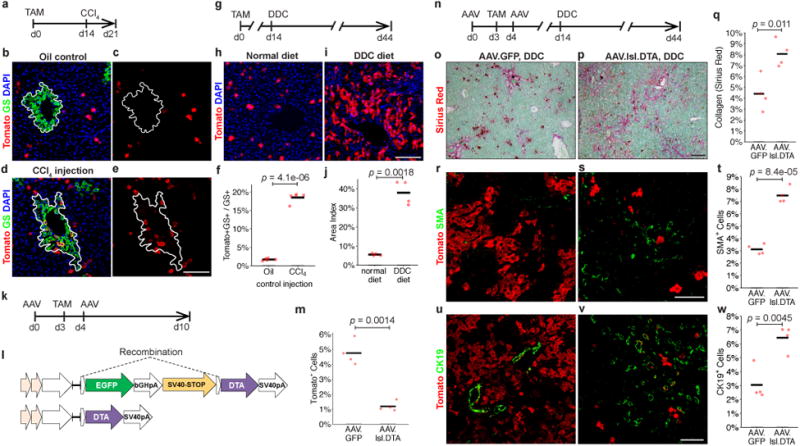
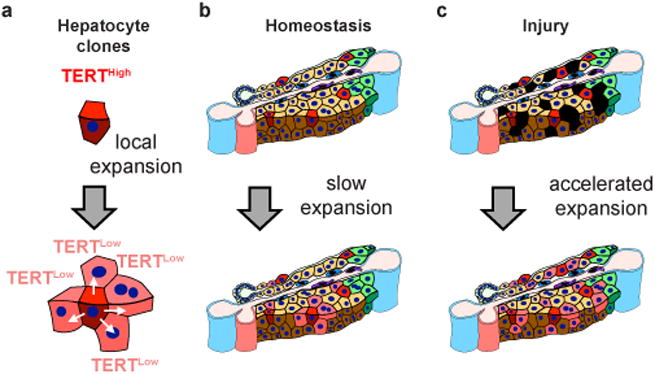
Hepatocytes are replenished gradually during homeostasis and robustly after liver injury1, 2. In adults, new hepatocytes originate from the existing hepatocyte pool3-8, but the cellular source of renewing hepatocytes remains unclear. Telomerase is expressed in many stem cell populations, and mutations in telomerase pathway genes have been linked to liver diseases9-11. Here we identify a subset of hepatocytes that expresses high levels of telomerase and show that this hepatocyte subset repopulates the liver during homeostasis and injury. Using lineage tracing from the telomerase reverse transcriptase (Tert) locus in mice, we demonstrate that rare hepatocytes with high telomerase expression (TERTHigh hepatocytes) are distributed throughout the liver lobule. During homeostasis, these cells regenerate hepatocytes in all lobular zones, and both self-renew and differentiate to yield expanding hepatocyte clones that eventually dominate the liver. In response to injury, the repopulating activity of TERTHigh hepatocytes is accelerated and their progeny cross zonal boundaries. RNA sequencing shows that metabolic genes are downregulated in TERTHigh hepatocytes, indicating that metabolic activity and repopulating activity may be segregated within the hepatocyte lineage. Genetic ablation of TERTHigh hepatocytes combined with chemical injury causes a marked increase in stellate cell activation and fibrosis. These results provide support for a 'distributed model' of hepatocyte renewal in which a subset of hepatocytes dispersed throughout the lobule clonally expands to maintain liver mass.
Figures
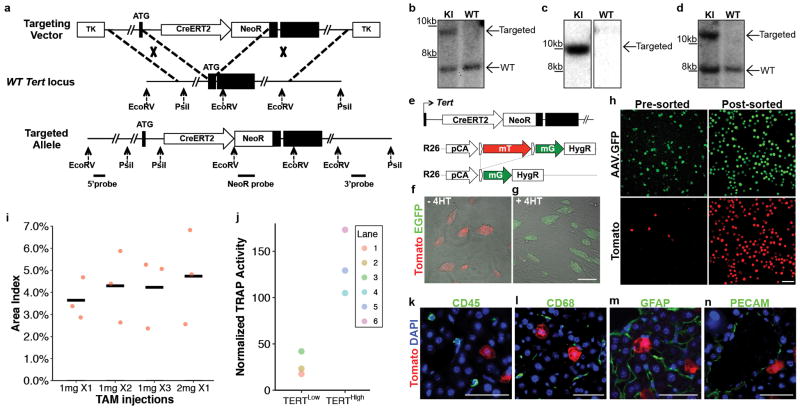
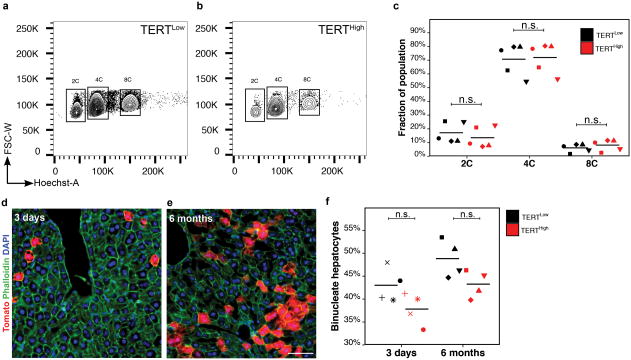
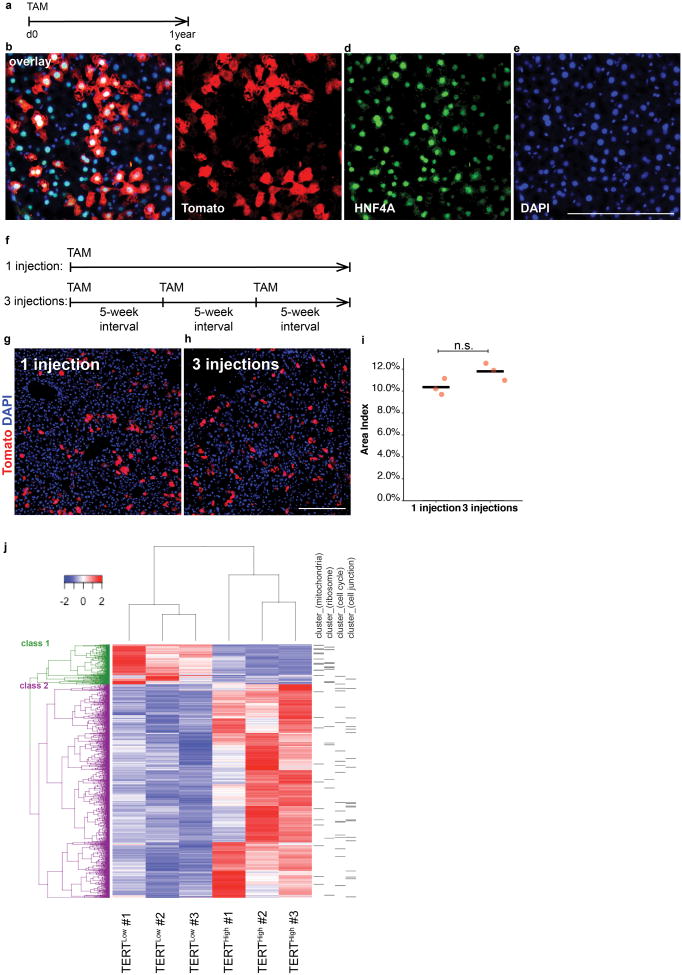
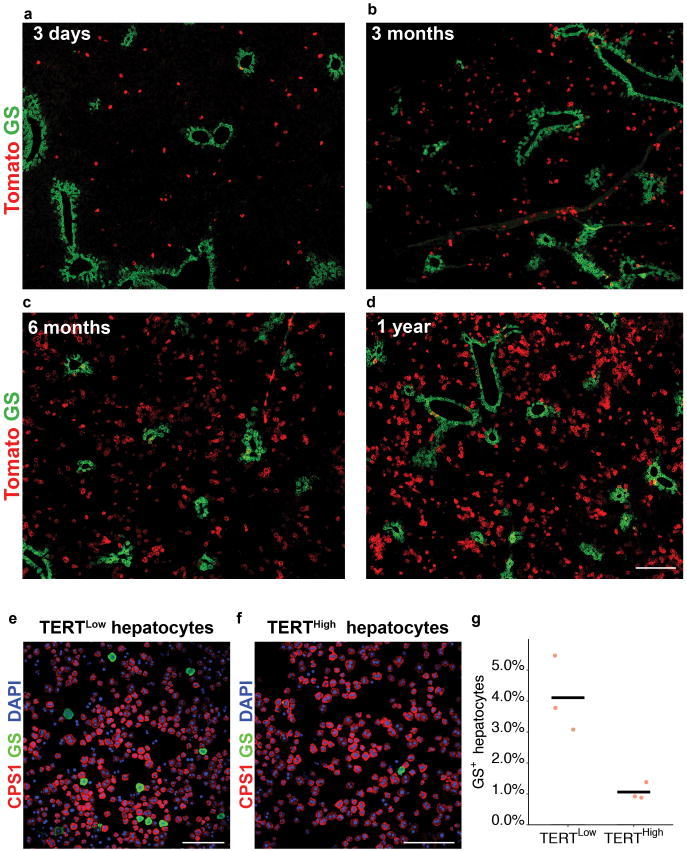
Comment in
-
The telomerase enzyme and liver renewal.Nature. 2018 Apr;556(7700):181-182. doi: 10.1038/d41586-018-02684-w. Nature. 2018. PMID: 29636575 No abstract available.
-
Replenishing hepatocytes.Nat Rev Gastroenterol Hepatol. 2018 Jun;15(6):328. doi: 10.1038/s41575-018-0017-6. Nat Rev Gastroenterol Hepatol. 2018. PMID: 29670251 No abstract available.
-
Telomerase Activity Links to Regenerative Capacity of Hepatocytes.Transplantation. 2018 Oct;102(10):1587-1588. doi: 10.1097/TP.0000000000002346. Transplantation. 2018. PMID: 30247437 No abstract available.
-
Highlight report: liver regeneration by a subset of hepatocytes with high expression of telomerase.Arch Toxicol. 2019 Dec;93(12):3633-3634. doi: 10.1007/s00204-019-02608-1. Epub 2019 Nov 1. Arch Toxicol. 2019. PMID: 31677075 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
