

Morphogen gradient reconstitution reveals Hedgehog pathway design principles
- PMID: 29622726
- PMCID: PMC6516753
- DOI: 10.1126/science.aao0645
Morphogen gradient reconstitution reveals Hedgehog pathway design principles
Abstract
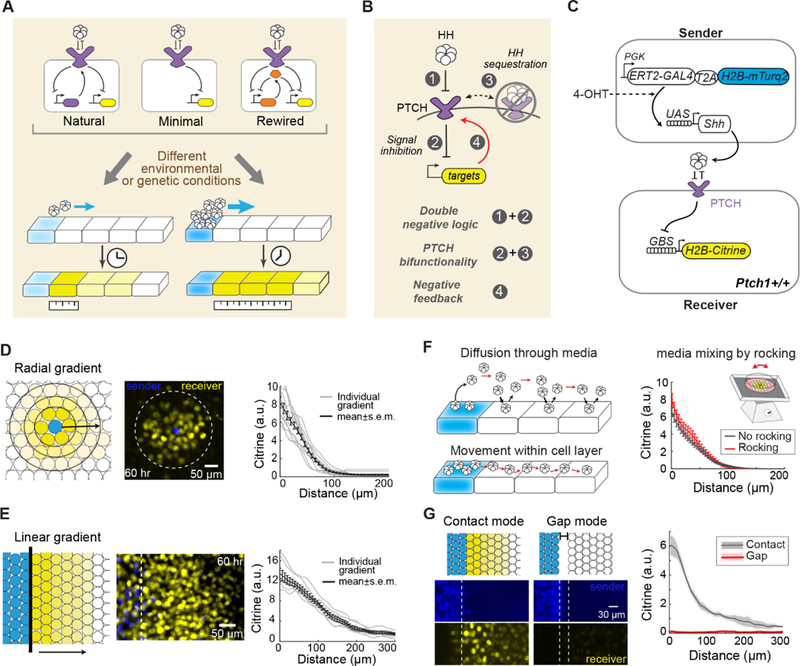
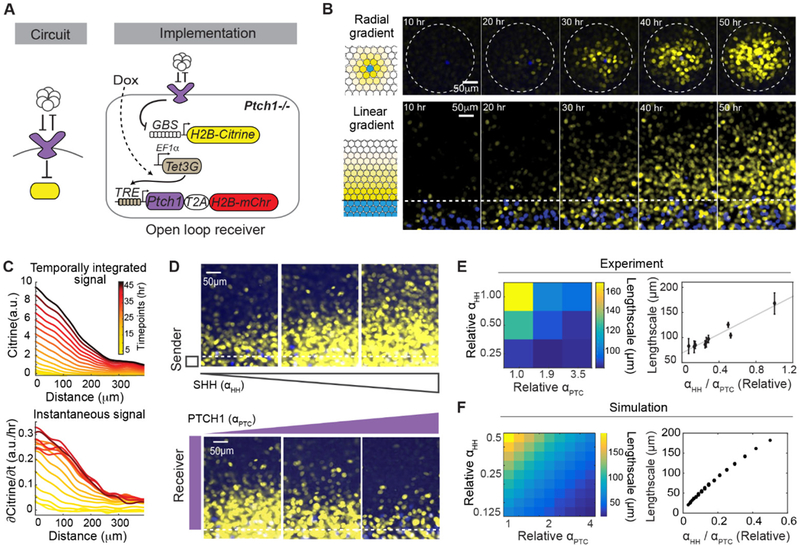
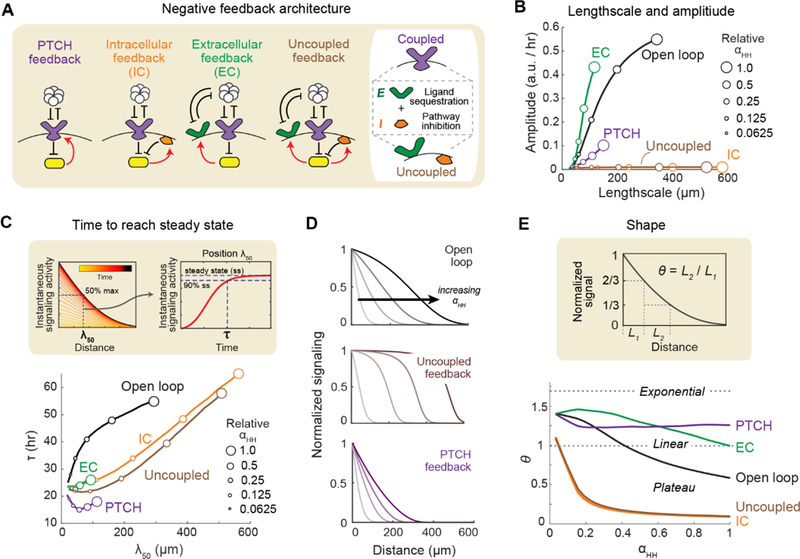
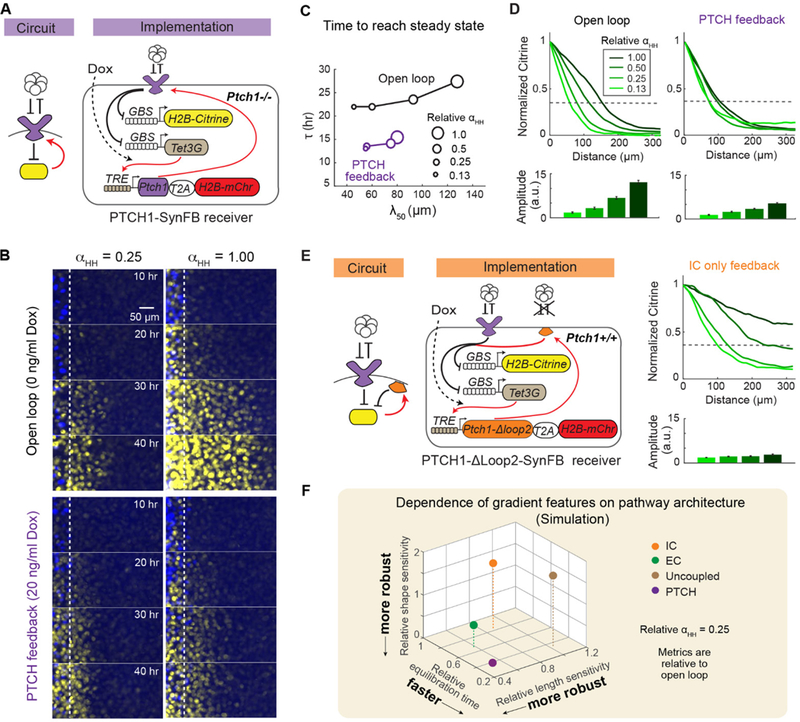
In developing tissues, cells estimate their spatial position by sensing graded concentrations of diffusible signaling proteins called morphogens. Morphogen-sensing pathways exhibit diverse molecular architectures, whose roles in controlling patterning dynamics and precision have been unclear. In this work, combining cell-based in vitro gradient reconstitution, genetic rewiring, and mathematical modeling, we systematically analyzed the distinctive architectural features of the Sonic Hedgehog pathway. We found that the combination of double-negative regulatory logic and negative feedback through the PTCH receptor accelerates gradient formation and improves robustness to variation in the morphogen production rate compared with alternative designs. The ability to isolate morphogen patterning from concurrent developmental processes and to compare the patterning behaviors of alternative, rewired pathway architectures offers a powerful way to understand and engineer multicellular patterning.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Comment in
-
Developmental Biology: Morphogen in a Dish.Curr Biol. 2018 Jul 9;28(13):R755-R757. doi: 10.1016/j.cub.2018.05.047. Curr Biol. 2018. PMID: 29990461
References
-
- Chen Y, Struhl G, Dual roles for patched in sequestering and transducing Hedgehog. Cell 87, 553–563 (1996). doi:1Q.1Q16/S0092-8674(00)81374-4 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
