

Influenza Infection in Humans Induces Broadly Cross-Reactive and Protective Neuraminidase-Reactive Antibodies
- PMID: 29625056
- PMCID: PMC5890936
- DOI: 10.1016/j.cell.2018.03.030
Influenza Infection in Humans Induces Broadly Cross-Reactive and Protective Neuraminidase-Reactive Antibodies
Abstract
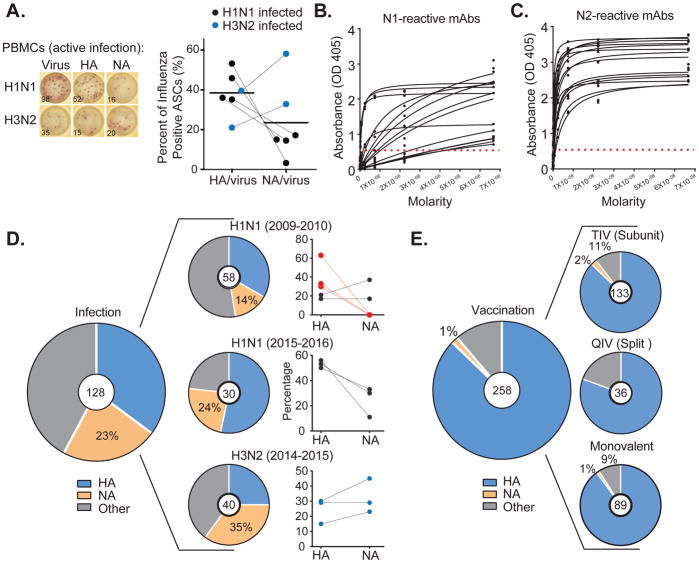
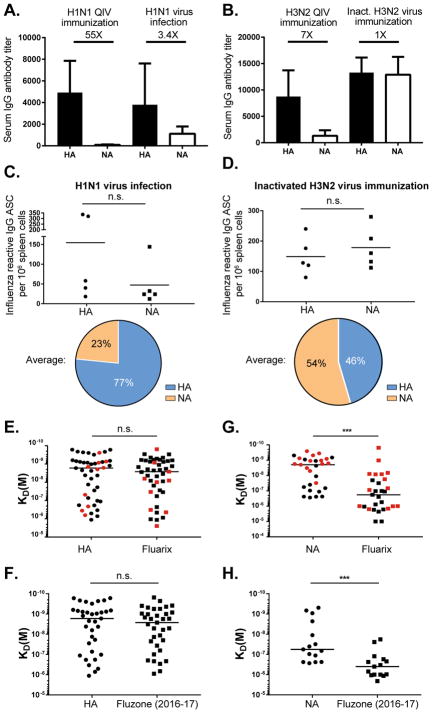
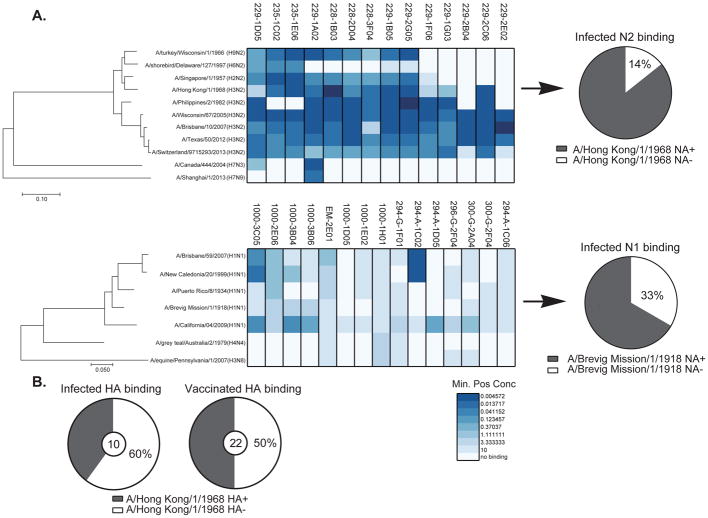
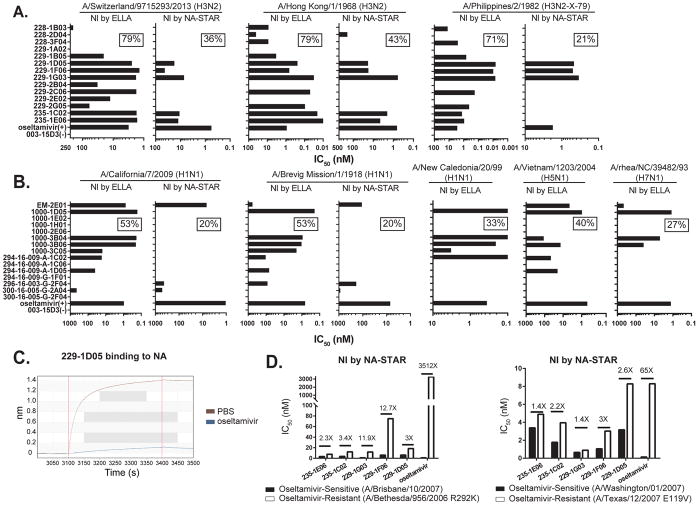
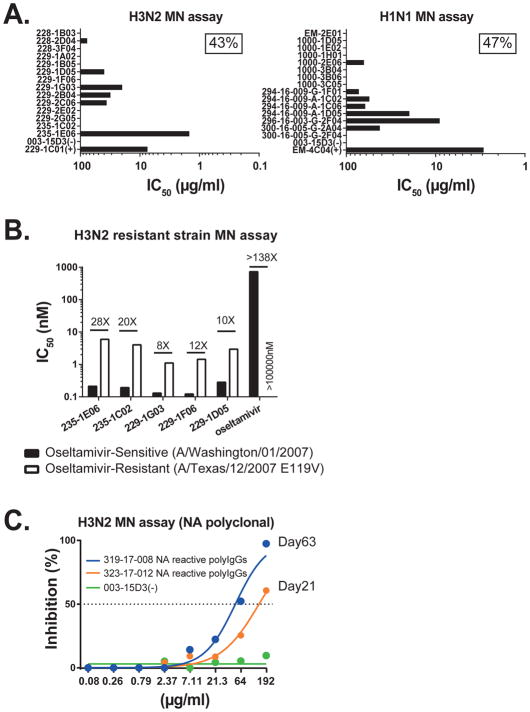
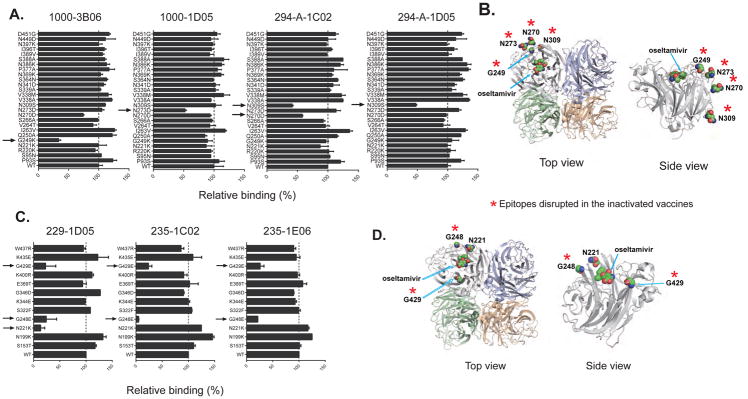
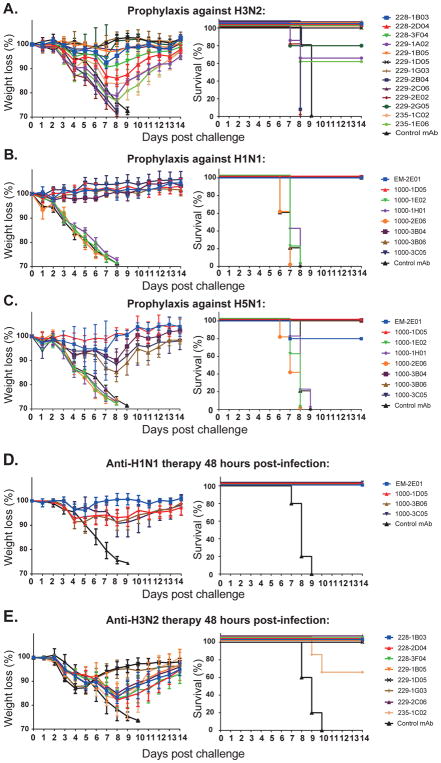
Antibodies to the hemagglutinin (HA) and neuraminidase (NA) glycoproteins are the major mediators of protection against influenza virus infection. Here, we report that current influenza vaccines poorly display key NA epitopes and rarely induce NA-reactive B cells. Conversely, influenza virus infection induces NA-reactive B cells at a frequency that approaches (H1N1) or exceeds (H3N2) that of HA-reactive B cells. NA-reactive antibodies display broad binding activity spanning the entire history of influenza A virus circulation in humans, including the original pandemic strains of both H1N1 and H3N2 subtypes. The antibodies robustly inhibit the enzymatic activity of NA, including oseltamivir-resistant variants, and provide robust prophylactic protection, including against avian H5N1 viruses, in vivo. When used therapeutically, NA-reactive antibodies protected mice from lethal influenza virus challenge even 48 hr post infection. These findings strongly suggest that influenza vaccines should be optimized to improve targeting of NA for durable and broad protection against divergent influenza strains.
Keywords: B cell; human immunology; humoral immune response; influenza; monoclonal antibody; neuraminidase; therapeutics; vaccine; virus infection.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Influenza virus: Choose your enemies wisely.Nat Rev Immunol. 2018 Apr 25;18(5):294-295. doi: 10.1038/nri.2018.30. Nat Rev Immunol. 2018. PMID: 29692419 No abstract available.
References
-
- Abed Y, Hardy I, Li Y, Boivin G. Divergent evolution of hemagglutinin and neuraminidase genes in recent influenza A:H3N2 viruses isolated in Canada. J Med Virol. 2002;67:589–595. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
