

KLRG1+ Effector CD8+ T Cells Lose KLRG1, Differentiate into All Memory T Cell Lineages, and Convey Enhanced Protective Immunity
- PMID: 29625895
- PMCID: PMC6465538
- DOI: 10.1016/j.immuni.2018.03.015
KLRG1+ Effector CD8+ T Cells Lose KLRG1, Differentiate into All Memory T Cell Lineages, and Convey Enhanced Protective Immunity
Abstract
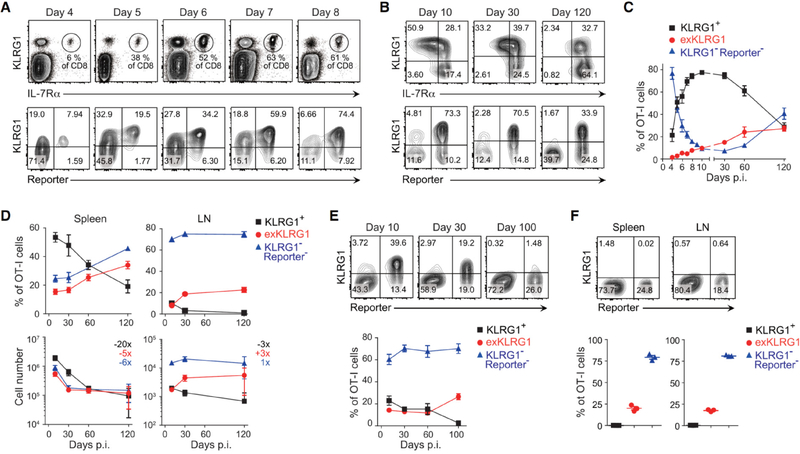
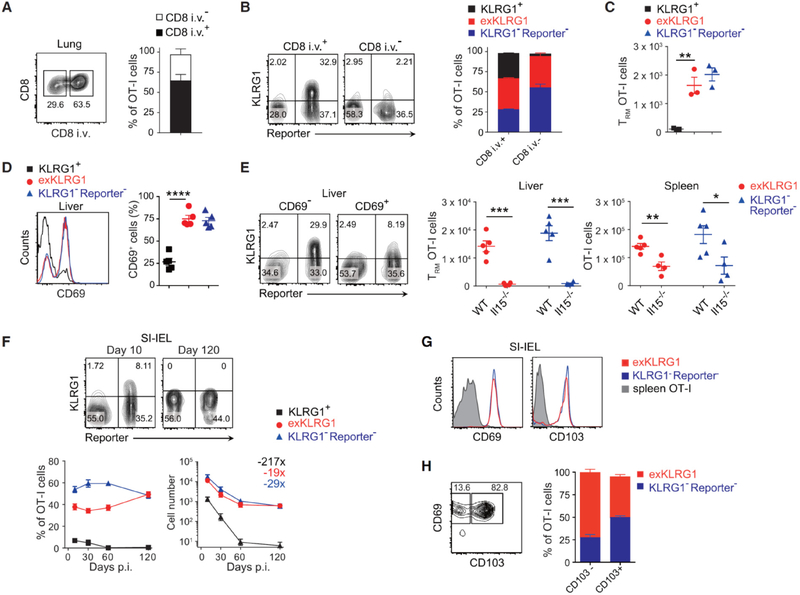
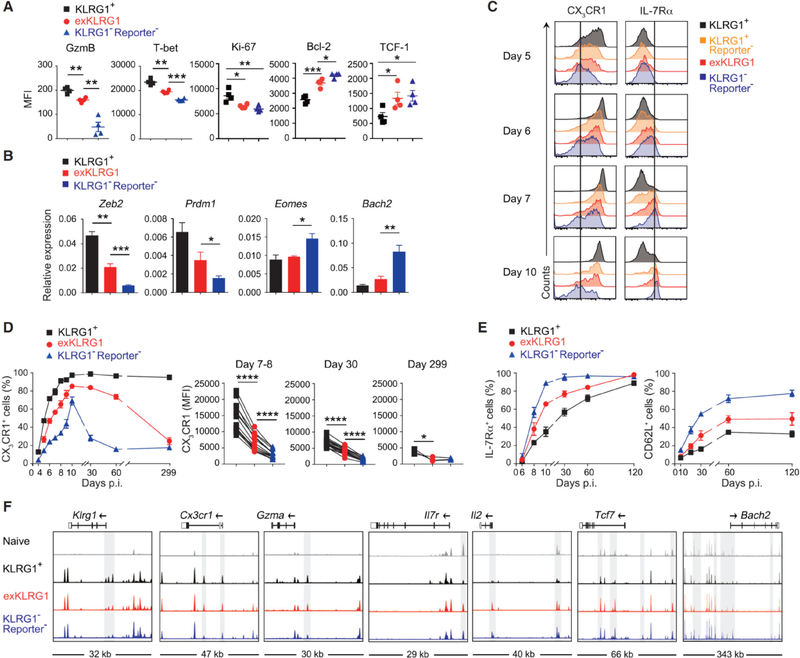
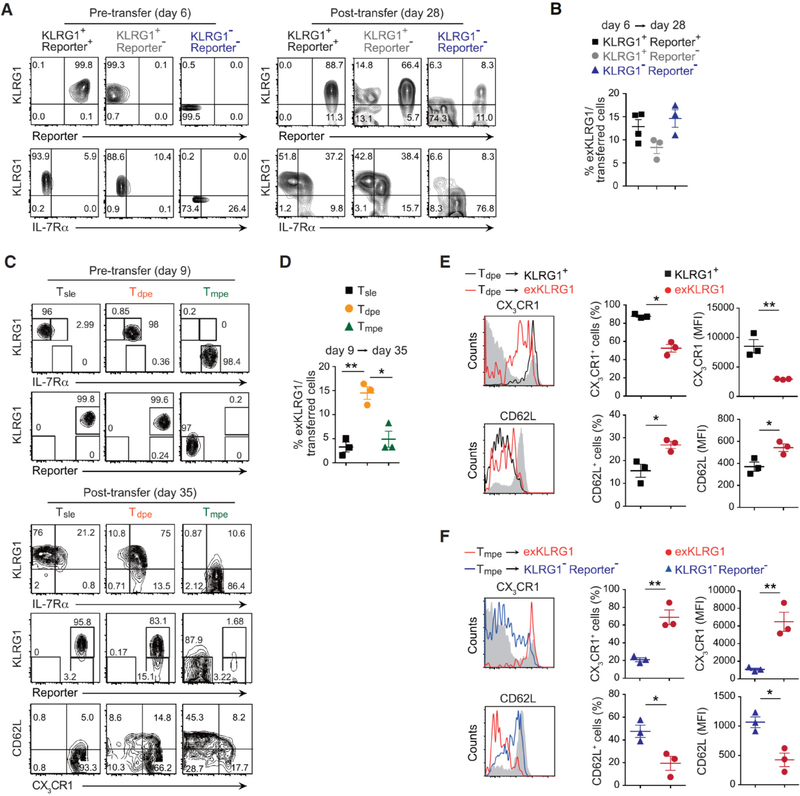
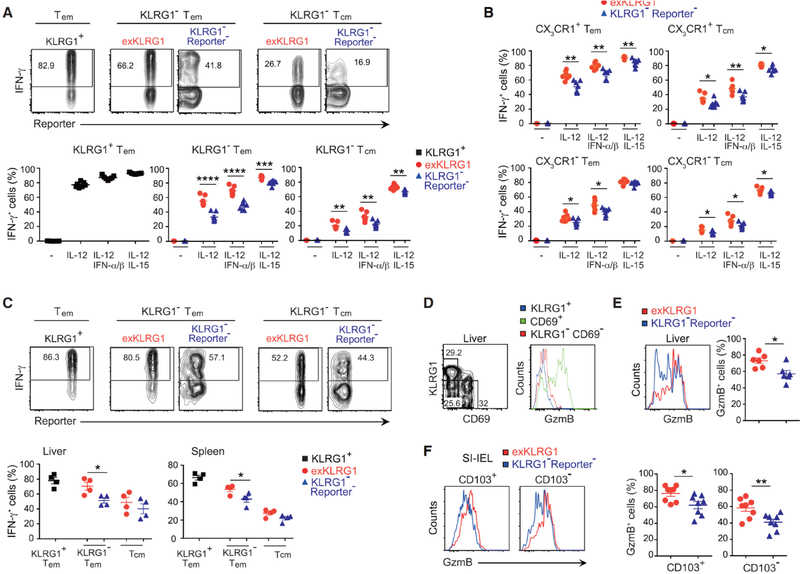
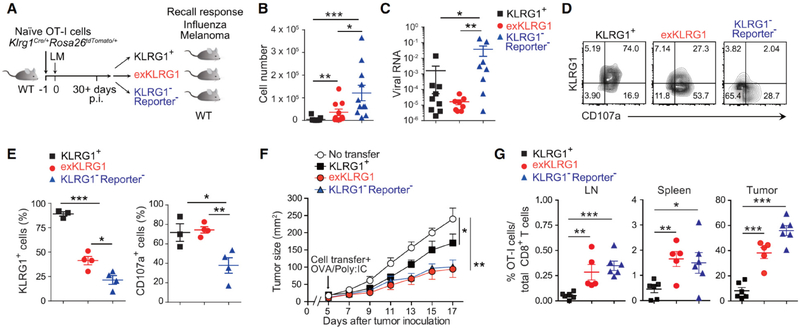
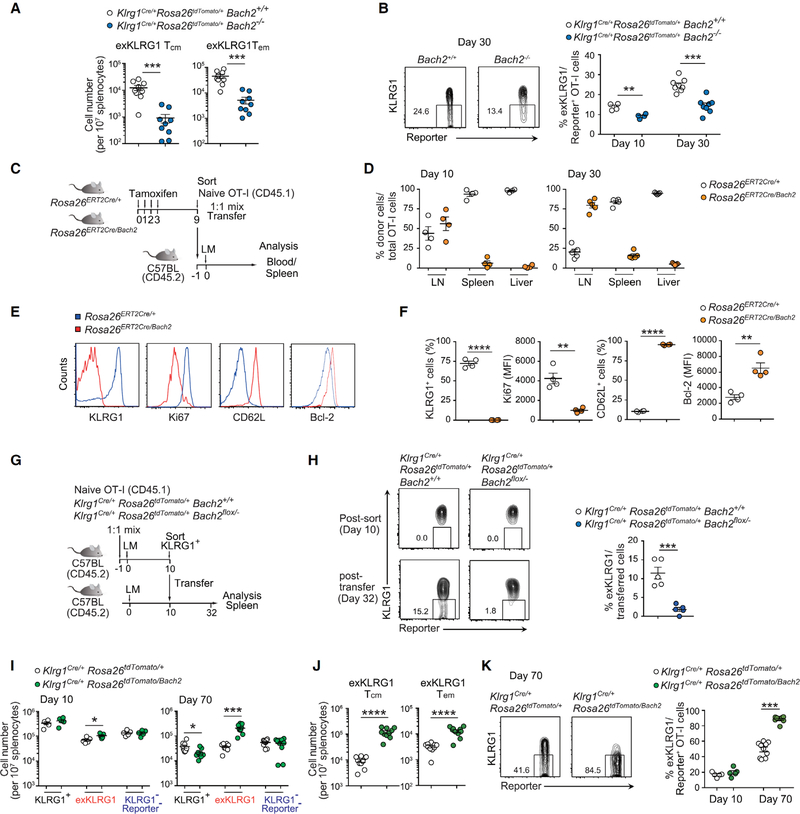
Protective immunity against pathogens depends on the efficient generation of functionally diverse effector and memory T lymphocytes. However, whether plasticity during effector-to-memory CD8+ T cell differentiation affects memory lineage specification and functional versatility remains unclear. Using genetic fate mapping analysis of highly cytotoxic KLRG1+ effector CD8+ T cells, we demonstrated that KLRG1+ cells receiving intermediate amounts of activating and inflammatory signals downregulated KLRG1 during the contraction phase in a Bach2-dependent manner and differentiated into all memory T cell linages, including CX3CR1int peripheral memory cells and tissue-resident memory cells. "ExKLRG1" memory cells retained high cytotoxic and proliferative capacity distinct from other populations, which contributed to effective anti-influenza and anti-tumor immunity. Our work demonstrates that developmental plasticity of KLRG1+ effector CD8+ T cells is important in promoting functionally versatile memory cells and long-term protective immunity.
Keywords: Bach2; CD8 T cell; CX(3)CR1; cancer; fate mapping; inflammation; influenza; memory; plasticity; tissue-resident.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Comment in
-
Bach2: An Instrument of Heterogeneity for Long-Term Protection.Immunity. 2018 Apr 17;48(4):618-620. doi: 10.1016/j.immuni.2018.03.033. Immunity. 2018. PMID: 29669243
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
