

A Y-Encoded Suppressor of Feminization Arose via Lineage-Specific Duplication of a Cytokinin Response Regulator in Kiwifruit
- PMID: 29626069
- PMCID: PMC5969274
- DOI: 10.1105/tpc.17.00787
A Y-Encoded Suppressor of Feminization Arose via Lineage-Specific Duplication of a Cytokinin Response Regulator in Kiwifruit
Abstract
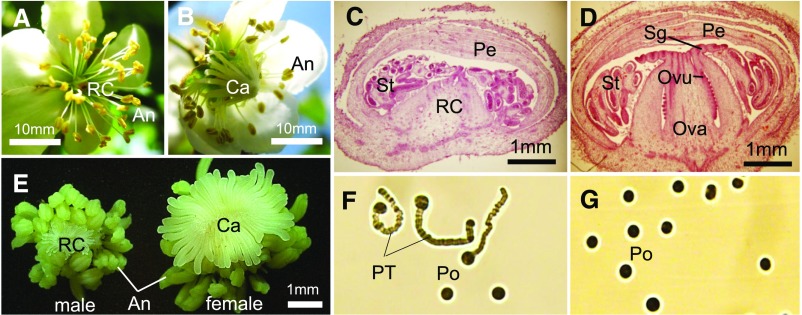
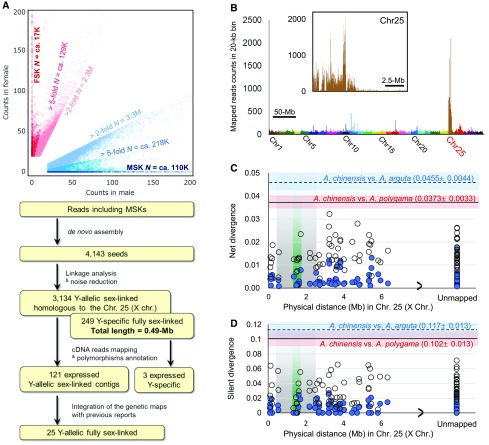
Dioecy, the presence of male and female flowers on distinct individuals, has evolved independently in multiple plant lineages, and the genes involved in this differential development are just starting to be uncovered in a few species. Here, we used genomic approaches to investigate this pathway in kiwifruits (genus Actinidia). Genome-wide cataloging of male-specific subsequences, combined with transcriptome analysis, led to the identification of a type-C cytokinin response regulator as a potential sex determinant gene in this genus. Functional transgenic analyses in two model systems, Arabidopsis thaliana and Nicotiana tabacum, indicated that this gene acts as a dominant suppressor of carpel development, prompting us to name it Shy Girl (SyGI). Evolutionary analyses in a panel of Actinidia species revealed that SyGI is located in the Y-specific region of the genome and probably arose from a lineage-specific gene duplication. Comparisons with the duplicated autosomal counterpart, and with orthologs from other angiosperms, suggest that the SyGI-specific duplication and subsequent evolution of cis-elements may have played a key role in the acquisition of separate sexes in this species.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures






Comment in
-
Shy Girl Gives Kiwifruit Male Flowers.Plant Cell. 2018 Apr;30(4):739-740. doi: 10.1105/tpc.18.00278. Epub 2018 Apr 6. Plant Cell. 2018. PMID: 29626070 Free PMC article. No abstract available.
References
-
- Akagi T., Henry I.M., Tao R., Comai L. (2014). Plant genetics. A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 346: 646–650. - PubMed
-
- Argueso C.T., Raines T., Kieber J.J. (2010). Cytokinin signaling and transcriptional networks. Curr. Opin. Plant Biol. 13: 533–539. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
