

Sorting cells alters their redox state and cellular metabolome
- PMID: 29627745
- PMCID: PMC5952879
- DOI: 10.1016/j.redox.2018.03.004
Sorting cells alters their redox state and cellular metabolome
Abstract
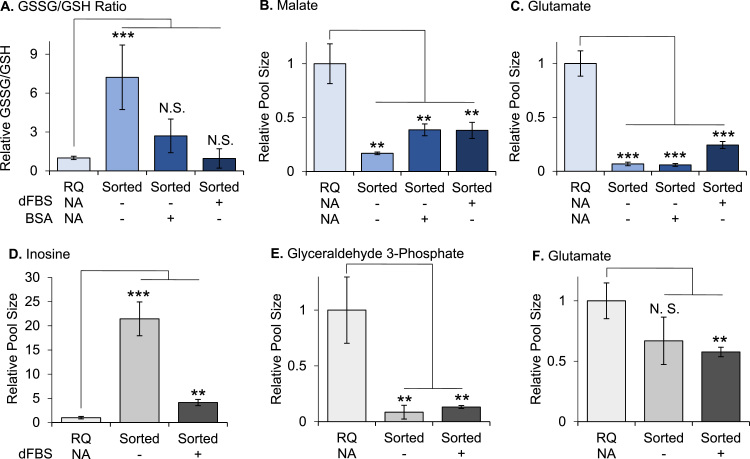
A growing appreciation of the metabolic artifacts of cell culture has generated heightened enthusiasm for performing metabolomics on populations of cells purified from tissues and biofluids. Fluorescence activated cell sorting, or FACS, is a widely used experimental approach to purify specific cell types from complex heterogeneous samples. Here we show that FACS introduces oxidative stress and alters the metabolic state of cells. Compared to unsorted controls, astrocytes subjected to FACS prior to metabolomic analysis showed altered ratios of GSSG to GSH, NADPH to NADP+, and NAD+ to NADH. Additionally, a 50% increase in reactive oxygen species was observed in astrocytes subjected to FACS relative to unsorted controls. At a more comprehensive scale, nearly half of the metabolomic features that we profiled by liquid chromatography/mass spectrometry were changed by at least 1.5-fold in intensity due to cell sorting. Some specific metabolites identified to have significantly altered levels as a result of cell sorting included glycogen, nucleosides, amino acids, central carbon metabolites, and acylcarnitines. Although the addition of fetal bovine serum to the cell-sorting buffer decreased oxidative stress and attenuated changes in metabolite concentrations, fetal bovine serum did not preserve the metabolic state of the cells during FACS. We conclude that, irrespective of buffer components and data-normalization strategies we examined, metabolomic results from sorted cells do not accurately reflect physiological conditions prior to sorting.
Copyright © 2018 The Authors. Published by Elsevier B.V. All rights reserved.
Figures



References
-
- Villas-Bôas S.G., Bruheim P. Cold glycerol–saline: the promising quenching solution for accurate intracellular metabolite analysis of microbial cells. Anal. Biochem. 2007;370:87–97. - PubMed
-
- Mayers J.R., Vander Heiden M.G. Nature and nurture: what determines tumor metabolic phenotypes? Cancer Res. 2017;77:3131–3134. - PubMed
-
- Zanetti K.A., Mette E., Maruvada P., Milner J., Moore S.C., Nicastro H.L., Ross S.A., Sampson J.N., Verma M., L J.S. The future of metabolomic profiling in population-based research: opportunities and challenges. J. Anal. Bioanal. Tech. 2014;5:1–4.
-
- Ng P.C., Lam H.S. Biomarkers in neonatology: the next generation of tests. Neonatology. 2012;102:145–151. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
