

Myelination of Neuronal Cell Bodies when Myelin Supply Exceeds Axonal Demand
- PMID: 29628374
- PMCID: PMC5912901
- DOI: 10.1016/j.cub.2018.02.068
Myelination of Neuronal Cell Bodies when Myelin Supply Exceeds Axonal Demand
Abstract
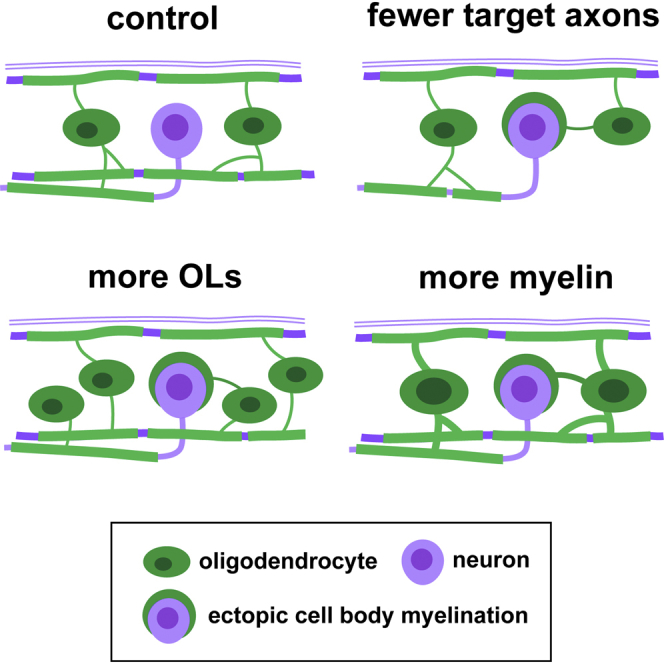
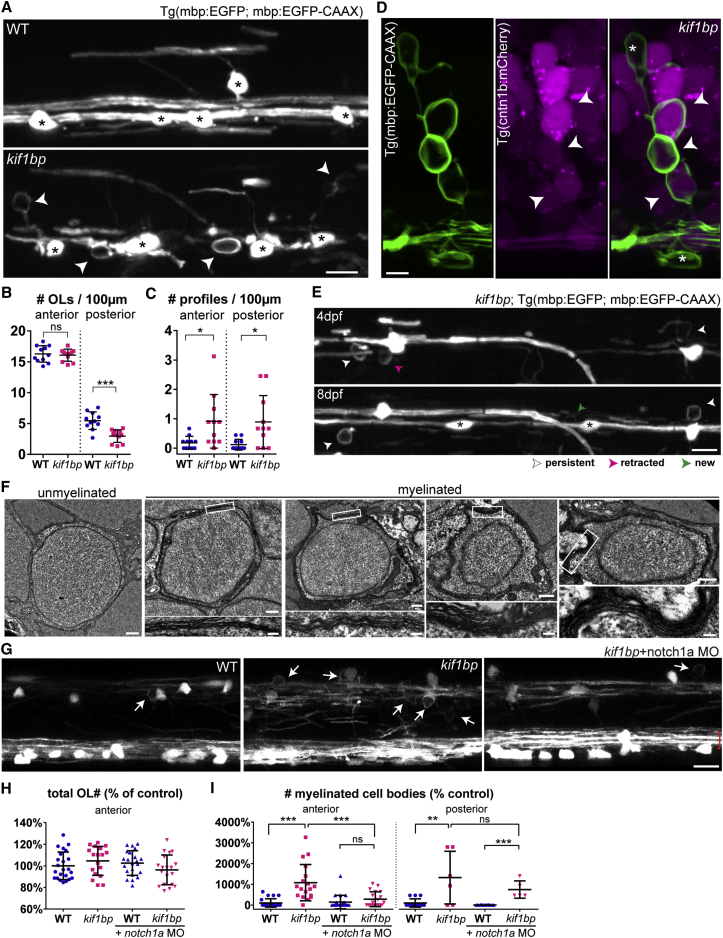
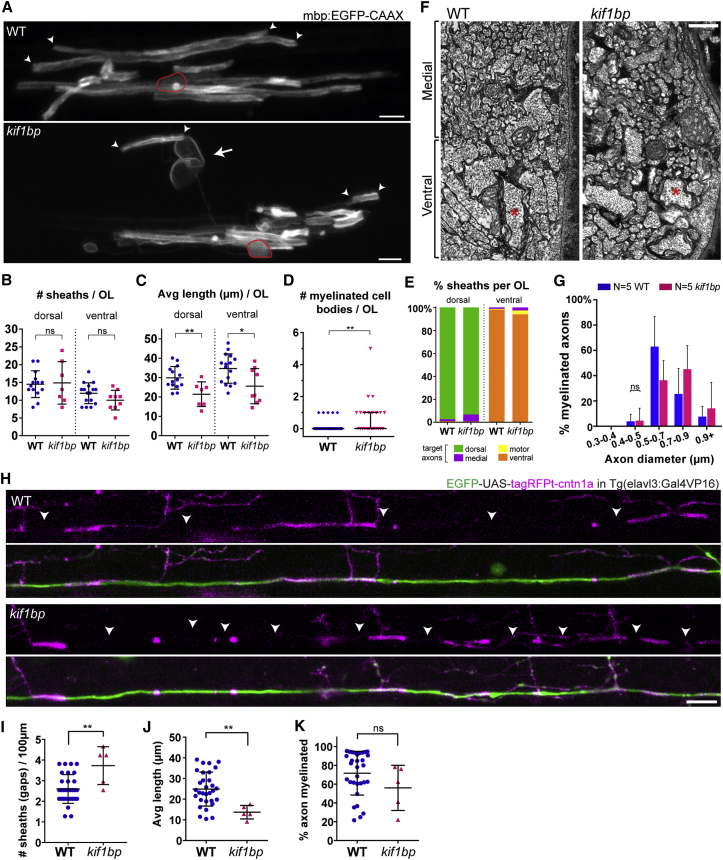
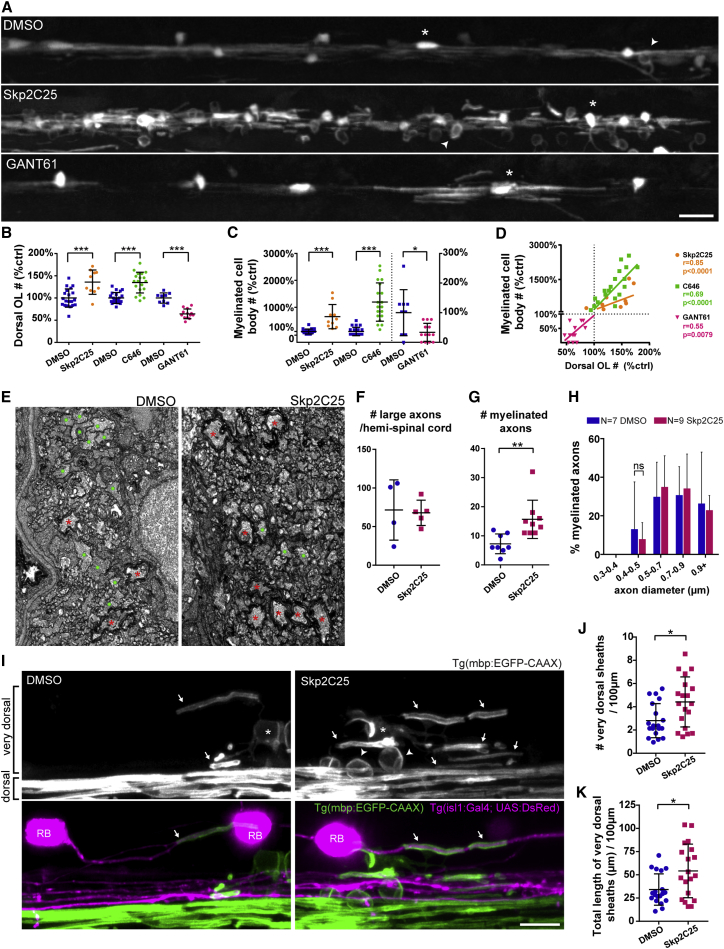
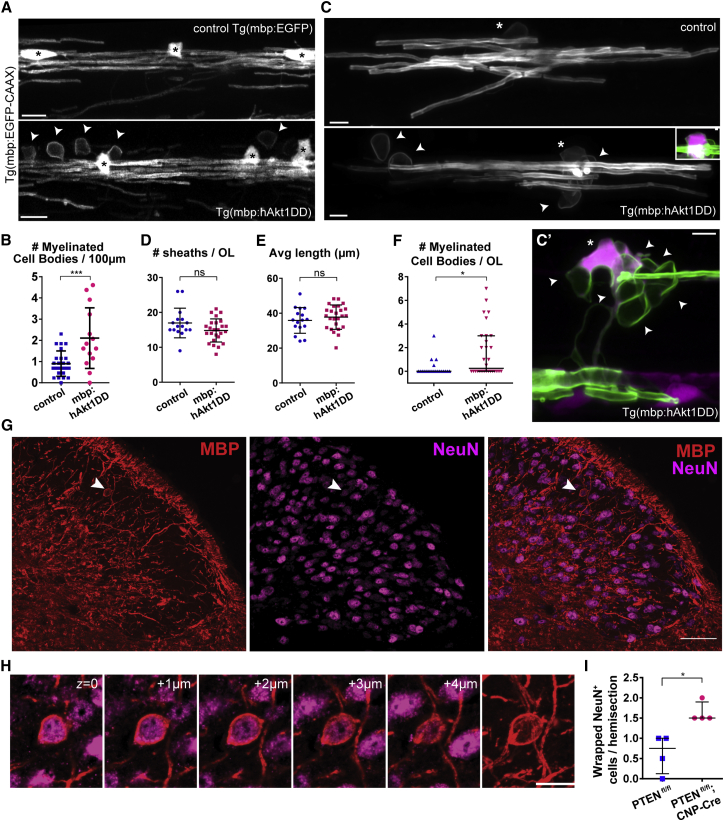
The correct targeting of myelin is essential for nervous system formation and function. Oligodendrocytes in the CNS myelinate some axons, but not others, and do not myelinate structures including cell bodies and dendrites [1]. Recent studies indicate that extrinsic signals, such as neuronal activity [2, 3] and cell adhesion molecules [4], can bias myelination toward some axons and away from cell bodies and dendrites, indicating that, in vivo, neuronal and axonal cues regulate myelin targeting. In vitro, however, oligodendrocytes have an intrinsic propensity to myelinate [5-7] and can promiscuously wrap inert synthetic structures resembling neuronal processes [8, 9] or cell bodies [4]. A current therapeutic goal for the treatment of demyelinating diseases is to greatly promote oligodendrogenesis [10-13]; thus, it is important to test how accurately extrinsic signals regulate the oligodendrocyte's intrinsic program of myelination in vivo. Here, we test the hypothesis that neurons regulate myelination with sufficient stringency to always ensure correct targeting. Surprisingly, however, we find that myelin targeting in vivo is not very stringent and that mistargeting occurs readily when oligodendrocyte and myelin supply exceed axonal demand. We find that myelin is mistargeted to neuronal cell bodies in zebrafish mutants with fewer axons and independently in drug-treated zebrafish with increased oligodendrogenesis. Additionally, by increasing myelin production of oligodendrocytes in zebrafish and mice, we find that excess myelin is also inappropriately targeted to cell bodies. Our results suggest that balancing oligodendrocyte-intrinsic programs of myelin supply with axonal demand is essential for correct myelin targeting in vivo and highlight potential liabilities of strongly promoting oligodendrogenesis.
Keywords: CNS; axon; myelin; myelin mistargeting; myelination; neuron; oligodendrocyte; zebrafish.
Copyright © 2018 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
