

Cancer metabolism: New insights into classic characteristics
- PMID: 29628997
- PMCID: PMC5884251
- DOI: 10.1016/j.jdsr.2017.08.003
Cancer metabolism: New insights into classic characteristics
Abstract
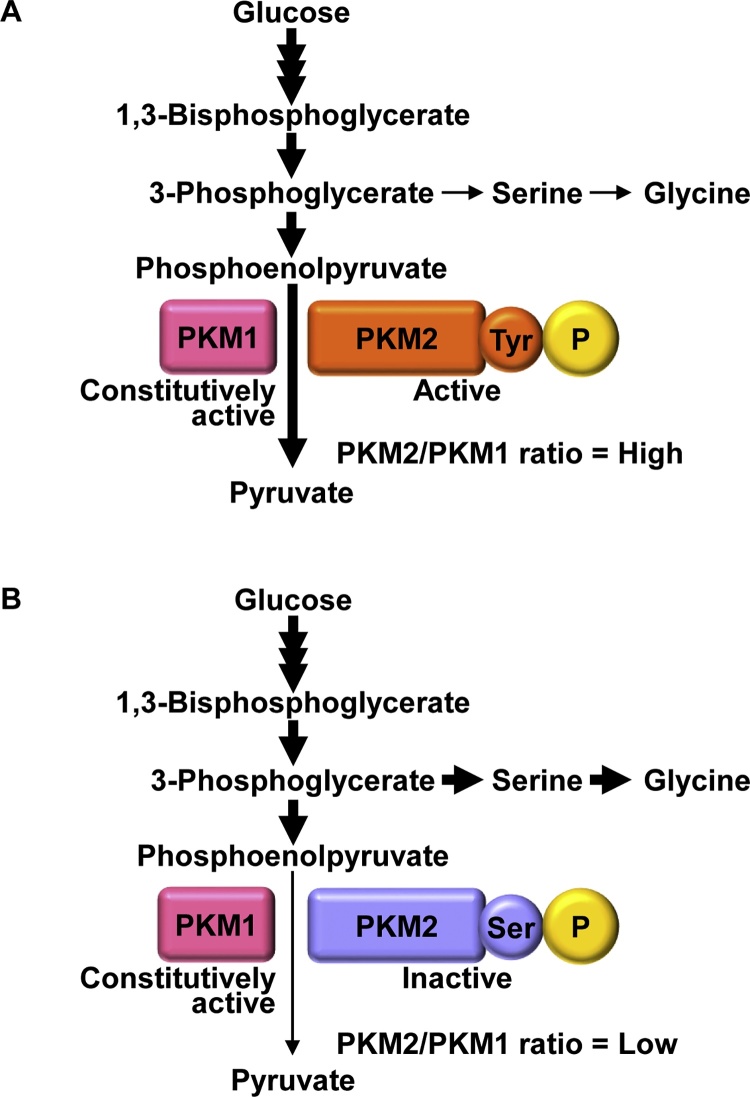
Initial studies of cancer metabolism in the early 1920s found that cancer cells were phenotypically characterized by aerobic glycolysis, in that these cells favor glucose uptake and lactate production, even in the presence of oxygen. This property, called the Warburg effect, is considered a hallmark of cancer. The mechanism by which these cells acquire aerobic glycolysis has been uncovered. Acidic extracellular fluid, secreted by cancer cells, induces a malignant phenotype, including invasion and metastasis. Cancer cells survival depends on a critical balance of redox status, which is regulated by amino acid metabolism. Glutamine is extremely important for oxidative phosphorylation and redox regulation. Cells highly dependent on glutamine and that cannot survive with glutamine are called glutamine-addicted cells. Metabolic reprogramming has been observed in cancer stem cells, which have the property of self-renewal and are resistant to chemotherapy and radiotherapy. These findings suggest that studies of cancer metabolism can reveal methods of preventing cancer recurrence and metastasis.
Keywords: Acidic extracellular pH; Glutamine metabolism; Glycolysis; Warburg effect.
Figures






References
-
- Warburg O., Posener K., Negelein E. Über den Stoffwechsel der Tumoren (On metabolism of tumors) Biochem Z. 1924;152:319–344.
-
- Warburg O. On the origin of cancer cells. Science. 1956;123(3191):309–314. - PubMed
-
- Engin K., Leeper D.B., Cater J.R., Thistlethwaite A.J., Tupchong L., McFarlane J.D. Extracellular pH distribution in human tumours. Int J Hyperthermia. 1995;11(2):211–216. - PubMed
-
- Delli Castelli D., Ferrauto G., Cutrin J.C., Terreno E., Aime S. In vivo maps of extracellular pH in murine melanoma by CEST-MRI. Magn Reson Med. 2014;71(1):326–332. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
