

E3 ubiquitin ligase SOR1 regulates ethylene response in rice root by modulating stability of Aux/IAA protein
- PMID: 29632179
- PMCID: PMC5924906
- DOI: 10.1073/pnas.1719387115
E3 ubiquitin ligase SOR1 regulates ethylene response in rice root by modulating stability of Aux/IAA protein
Abstract
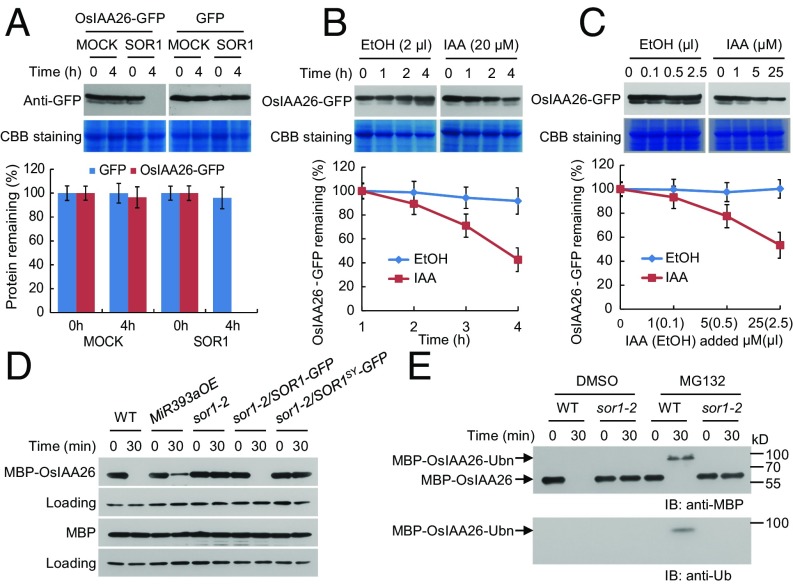
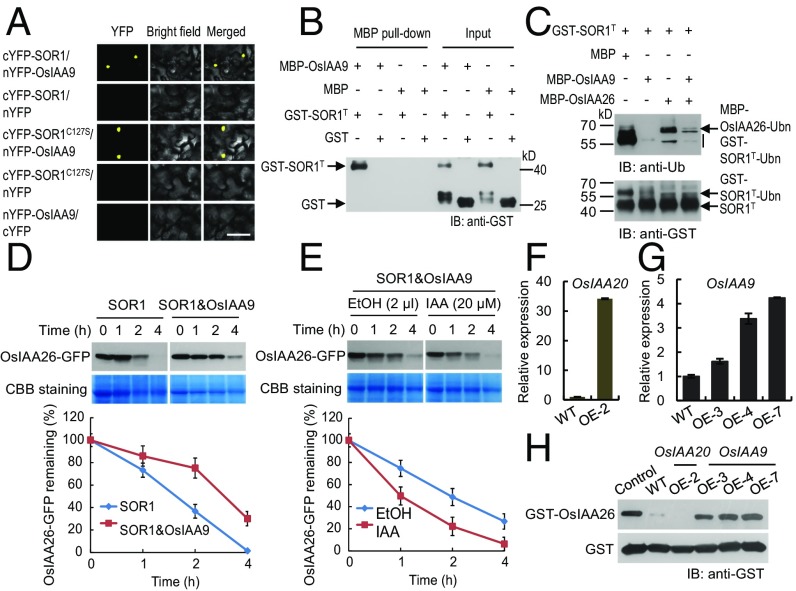
Plant hormones ethylene and auxin synergistically regulate plant root growth and development. Ubiquitin-mediated proteolysis of Aux/IAA transcriptional repressors by the E3 ubiquitin ligase SCFTIR1/AFB triggers a transcription-based auxin signaling. Here we show that rice (Oryza sativa L.) soil-surface rooting 1 (SOR1), which is a RING finger E3 ubiquitin ligase identified from analysis of a rice ethylene-insensitive mutant mhz2/sor1-2, controls root-specific ethylene responses by modulating Aux/IAA protein stability. SOR1 physically interacts with OsIAA26 and OsIAA9, which are atypical and canonical Aux/IAA proteins, respectively. SOR1 targets OsIAA26 for ubiquitin/26S proteasome-mediated degradation, whereas OsIAA9 protects the OsIAA26 protein from degradation by inhibiting the E3 activity of SOR1. Auxin promotes SOR1-dependent degradation of OsIAA26 by facilitating SCFOsTIR1/AFB2-mediated and SOR1-assisted destabilization of OsIAA9 protein. Our study provides a candidate mechanism by which the SOR1-OsIAA26 module acts downstream of the OsTIR1/AFB2-auxin-OsIAA9 signaling to modulate ethylene inhibition of root growth in rice seedlings.
Keywords: Aux/IAA protein; ethylene response; rice; root elongation; ubiquitination.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
