

Targeting the polyadenylation factor EhCFIm25 with RNA aptamers controls survival in Entamoeba histolytica
- PMID: 29632392
- PMCID: PMC5890266
- DOI: 10.1038/s41598-018-23997-w
Targeting the polyadenylation factor EhCFIm25 with RNA aptamers controls survival in Entamoeba histolytica
Abstract
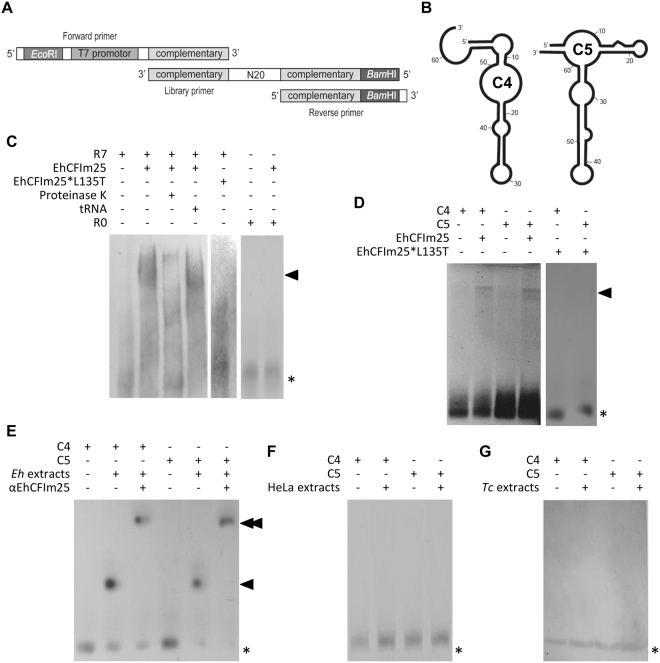
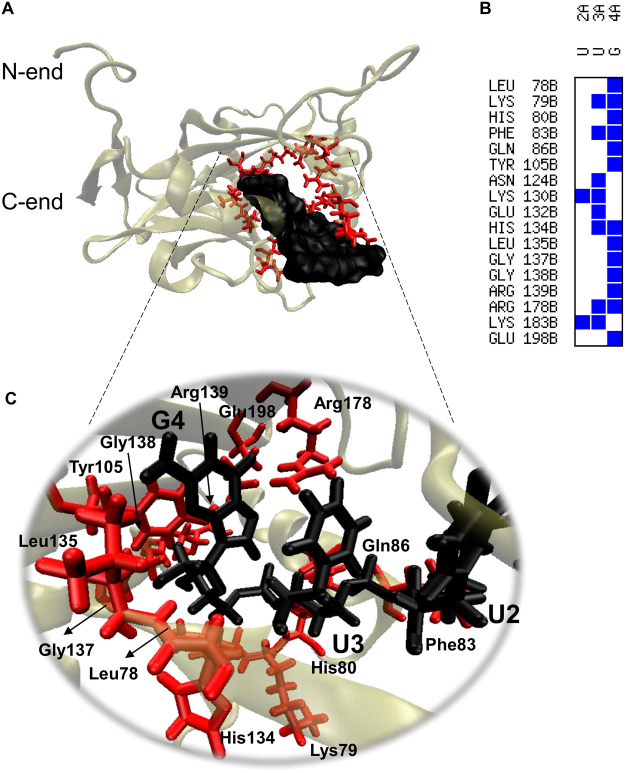
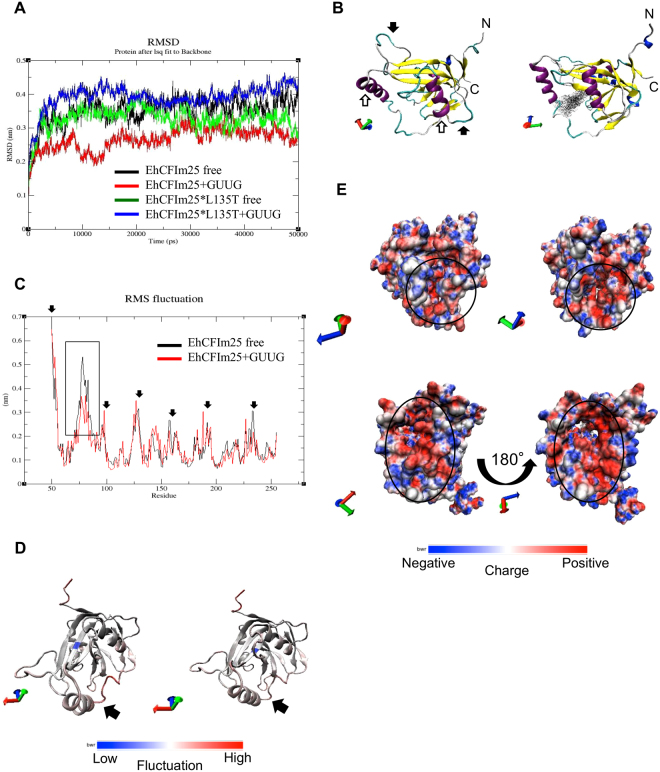
Messenger RNA 3'-end polyadenylation is an important regulator of gene expression in eukaryotic cells. In our search for new ways of treating parasitic infectious diseases, we looked at whether or not alterations in polyadenylation might control the survival of Entamoeba histolytica (the agent of amoebiasis in humans). We used molecular biology and computational tools to characterize the mRNA cleavage factor EhCFIm25, which is essential for polyadenylation in E. histolytica. By using a strategy based on the systematic evolution of ligands by exponential enrichment, we identified single-stranded RNA aptamers that target EhCFIm25. The results of RNA-protein binding assays showed that EhCFIm25 binds to the GUUG motif in vitro, which differs from the UGUA motif bound by the homologous human protein. Accordingly, docking experiments and molecular dynamic simulations confirmed that interaction with GUUG stabilizes EhCFIm25. Incubating E. histolytica trophozoites with selected aptamers inhibited parasite proliferation and rapidly led to cell death. Overall, our data indicate that targeting EhCFIm25 is an effective way of limiting the growth of E. histolytica in vitro. The present study is the first to have highlighted the potential value of RNA aptamers for controlling this human pathogen.
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Importance of amino acids Leu135 and Tyr236 for the interaction between EhCFIm25 and RNA: a molecular dynamics simulation study.J Mol Model. 2018 Jul 12;24(8):202. doi: 10.1007/s00894-018-3729-1. J Mol Model. 2018. PMID: 30003410
-
Silencing the cleavage factor CFIm25 as a new strategy to control Entamoeba histolytica parasite.J Microbiol. 2017 Oct;55(10):783-791. doi: 10.1007/s12275-017-7259-9. Epub 2017 Sep 28. J Microbiol. 2017. PMID: 28956353
-
Amino acid residues Leu135 and Tyr236 are required for RNA binding activity of CFIm25 in Entamoeba histolytica.Biochimie. 2015 Aug;115:44-51. doi: 10.1016/j.biochi.2015.04.017. Epub 2015 May 2. Biochimie. 2015. PMID: 25941172
-
Recent insights in pre-mRNA 3'-end processing signals and proteins in the protozoan parasite Entamoeba histolytica.Infect Disord Drug Targets. 2010 Aug;10(4):258-65. doi: 10.2174/187152610791591575. Infect Disord Drug Targets. 2010. PMID: 20429864 Review.
-
Life and Death of mRNA Molecules in Entamoeba histolytica.Front Cell Infect Microbiol. 2018 Jun 19;8:199. doi: 10.3389/fcimb.2018.00199. eCollection 2018. Front Cell Infect Microbiol. 2018. PMID: 29971219 Free PMC article. Review.
Cited by
-
Aptamer-based approaches for the detection of waterborne pathogens.Int Microbiol. 2021 May;24(2):125-140. doi: 10.1007/s10123-020-00154-w. Epub 2021 Jan 6. Int Microbiol. 2021. PMID: 33404933 Review.
-
Therapeutic applications of nucleic acid aptamers in microbial infections.J Biomed Sci. 2020 Jan 3;27(1):6. doi: 10.1186/s12929-019-0611-0. J Biomed Sci. 2020. PMID: 31900238 Free PMC article. Review.
-
Unraveling the relevance of the polyadenylation factor EhCFIm25 in Entamoeba histolytica through proteomic analysis.FEBS Open Bio. 2021 Oct;11(10):2819-2835. doi: 10.1002/2211-5463.13287. Epub 2021 Sep 13. FEBS Open Bio. 2021. PMID: 34486252 Free PMC article.
-
Current Advances in the Development of Diagnostic Tests based on Aptamers in Parasitology: A Systematic Review.Pharmaceutics. 2020 Oct 31;12(11):1046. doi: 10.3390/pharmaceutics12111046. Pharmaceutics. 2020. PMID: 33142793 Free PMC article. Review.
-
Recent Advances in Aptamer Discovery and Applications.Molecules. 2019 Mar 7;24(5):941. doi: 10.3390/molecules24050941. Molecules. 2019. PMID: 30866536 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
