

Continuous Change in Membrane and Membrane-Skeleton Organization During Development From Proerythroblast to Senescent Red Blood Cell
- PMID: 29632498
- PMCID: PMC5879444
- DOI: 10.3389/fphys.2018.00286
Continuous Change in Membrane and Membrane-Skeleton Organization During Development From Proerythroblast to Senescent Red Blood Cell
Abstract
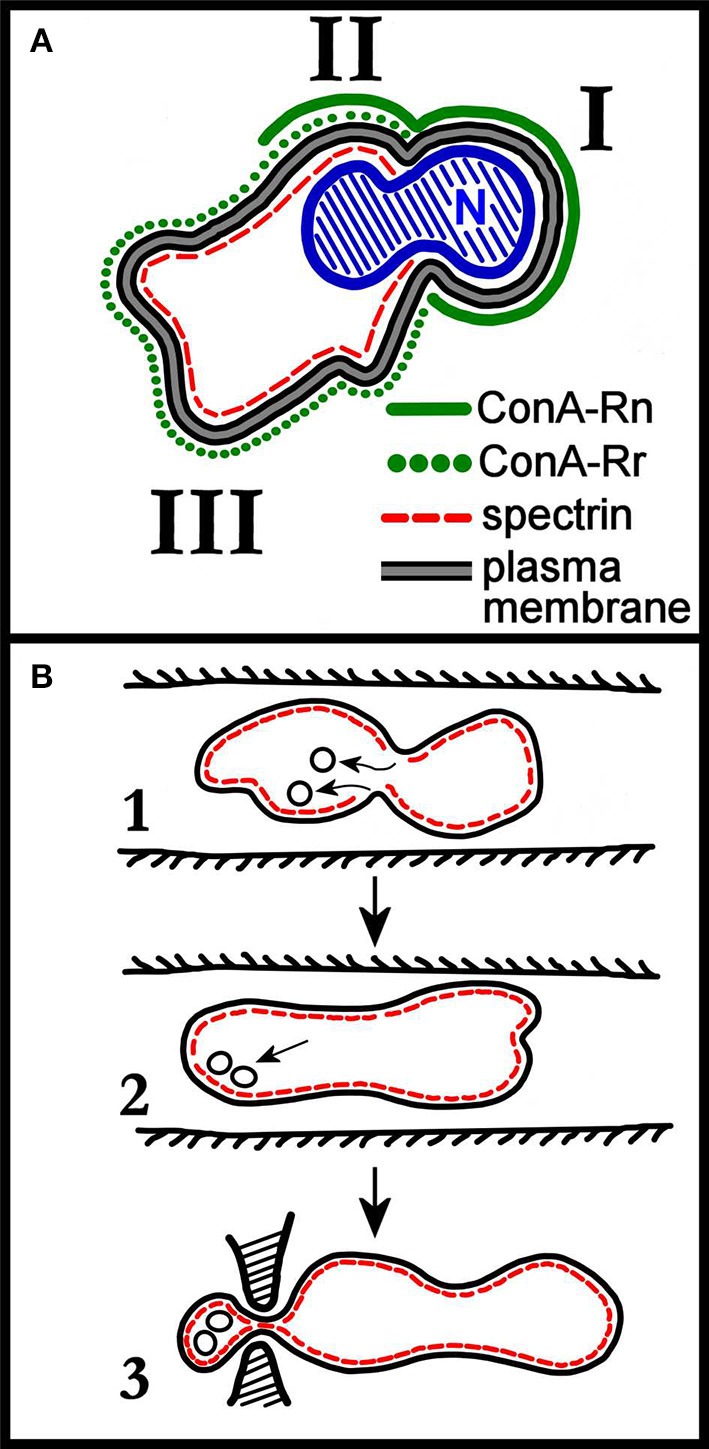
Within the context of erythropoiesis and the possibility of producing artificial red blood cells (RBCs) in vitro, a most critical step is the final differentiation of enucleated erythroblasts, or reticulocytes, to a fully mature biconcave discocyte, the RBC. Reviewed here is the current knowledge about this fundamental maturational process. By combining literature data with our own experimental evidence we propose that the early phase in the maturation of reticulocytes to RBCs is driven by a membrane raft-based mechanism for the sorting of disposable membrane proteins, mostly the no longer needed transferrin receptor (TfR), to the multivesicular endosome (MVE) as cargo of intraluminal vesicles that are subsequently exocytosed as exosomes, consistently with the seminal and original observation of Johnstone and collaborators of more than 30 years ago (Pan BT, Johnstone RM. Cell. 1983;33:967-978). According to a strikingly selective sorting process, the TfR becomes cargo destined to exocytosis while other molecules, including the most abundant RBC transmembrane protein, band 3, are completely retained in the cell membrane. It is also proposed that while this process could be operating in the early maturational steps in the bone marrow, additional mechanism(s) must be at play for the final removal of the excess reticulocyte membrane that is observed to occur in the circulation. This processing will most likely require the intervention of the spleen, whose function is also necessary for the continuous remodeling of the RBC membrane all along this cell's circulatory life.
Keywords: artificial red blood cells; autophagy; fluid phase endocytosis; membrane rafts; multivesicular endosome; red blood cell ageing; reticulocyte maturation; spleen.
Figures

References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources