

Variant-specific and reciprocal Hsp40 functions in Hsp104-mediated prion elimination
- PMID: 29633387
- PMCID: PMC6099457
- DOI: 10.1111/mmi.13966
Variant-specific and reciprocal Hsp40 functions in Hsp104-mediated prion elimination
Abstract
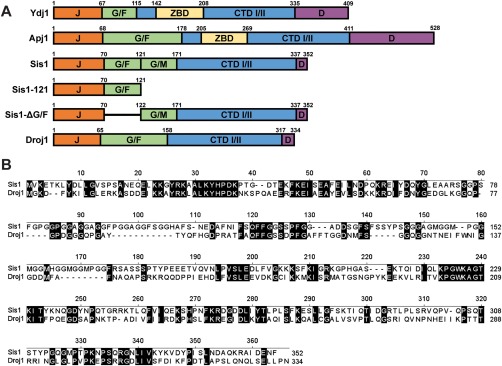
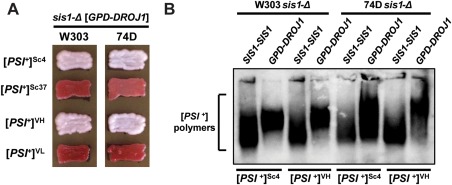
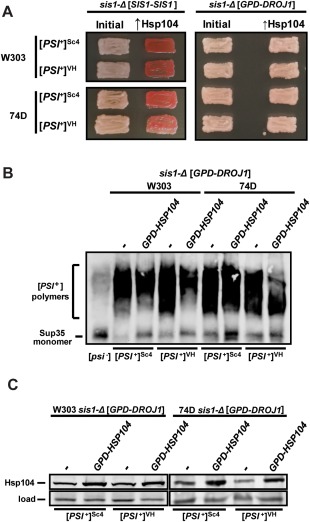
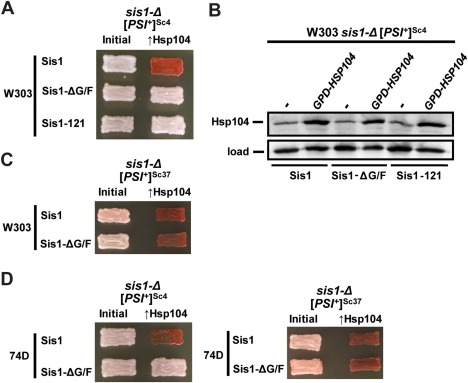
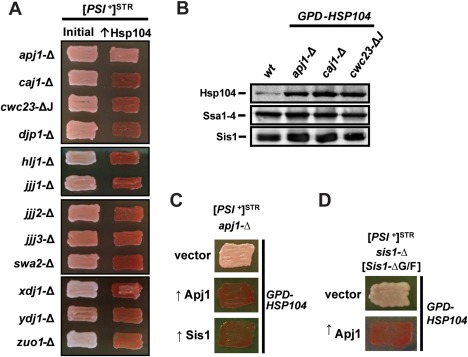
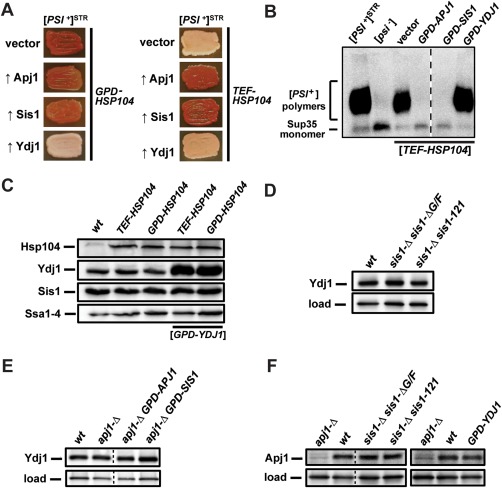
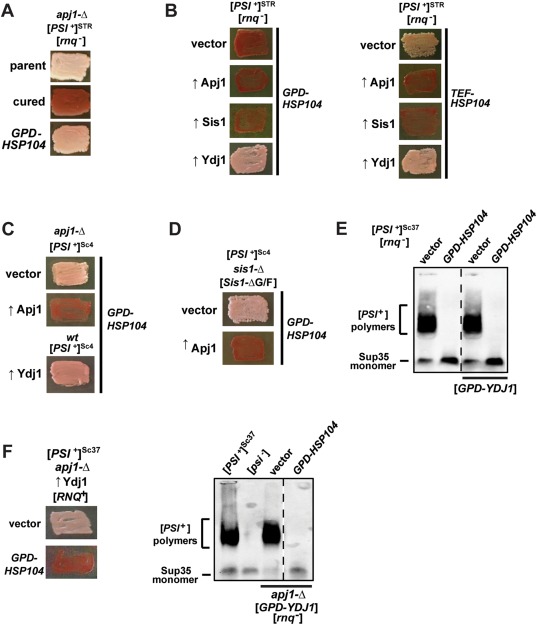
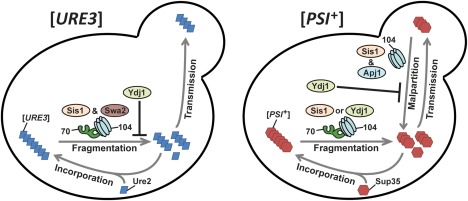
The amyloid-based prions of Saccharomyces cerevisiae are heritable aggregates of misfolded proteins, passed to daughter cells following fragmentation by molecular chaperones including the J-protein Sis1, Hsp70 and Hsp104. Overexpression of Hsp104 efficiently cures cell populations of the prion [PSI+ ] by an alternative Sis1-dependent mechanism that is currently the subject of significant debate. Here, we broadly investigate the role of J-proteins in this process by determining the impact of amyloid polymorphisms (prion variants) on the ability of well-studied Sis1 constructs to compensate for Sis1 and ask whether any other S. cerevisiae cytosolic J-proteins are also required for this process. Our comprehensive screen, examining all 13 members of the yeast cytosolic/nuclear J-protein complement, uncovered significant variant-dependent genetic evidence for a role of Apj1 (antiprion DnaJ) in this process. For strong, but not weak [PSI+ ] variants, depletion of Apj1 inhibits Hsp104-mediated curing. Overexpression of either Apj1 or Sis1 enhances curing, while overexpression of Ydj1 completely blocks it. We also demonstrated that Sis1 was the only J-protein necessary for the propagation of at least two weak [PSI+ ] variants and no J-protein alteration, or even combination of alterations, affected the curing of weak [PSI+ ] variants, suggesting the possibility of biochemically distinct, variant-specific Hsp104-mediated curing mechanisms.
© 2018 The Authors Molecular Microbiology Published by John Wiley & Sons Ltd.
Figures

References
-
- Allen, K.D. , Chernova, T.A. , Tennant, E.P. , Wilkinson, K.D. , and Chernoff, Y.O. (2007) Effects of ubiquitin system alterations on the formation and loss of a yeast prion. J Biol Chem 282: 3004–3013. - PubMed
-
- Bagriantsev, S.N. , Kushnirov, V.V. , and Liebman, S.W. (2006) Analysis of amyloid aggregates using agarose gel electrophoresis. Methods Enzymol 412: 33–48. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
