

TGFβ signaling related genes are involved in hormonal mediation during termite soldier differentiation
- PMID: 29641521
- PMCID: PMC5912798
- DOI: 10.1371/journal.pgen.1007338
TGFβ signaling related genes are involved in hormonal mediation during termite soldier differentiation
Abstract
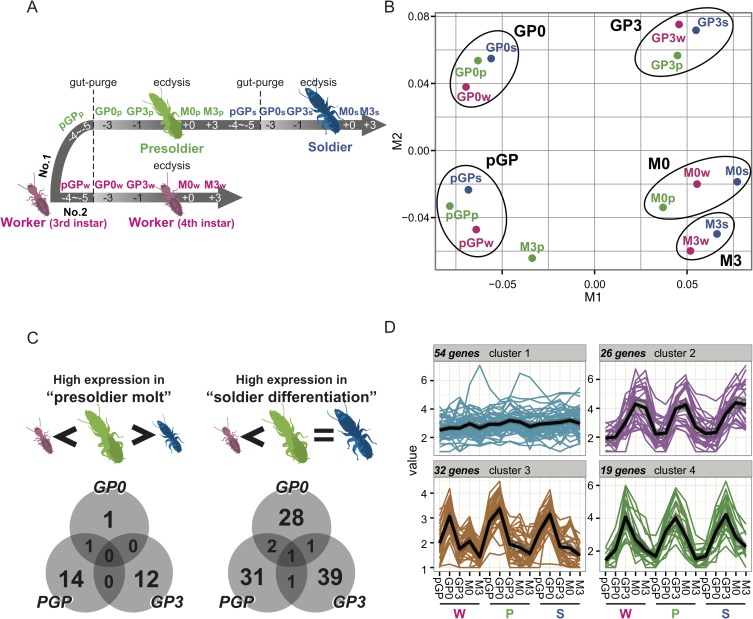
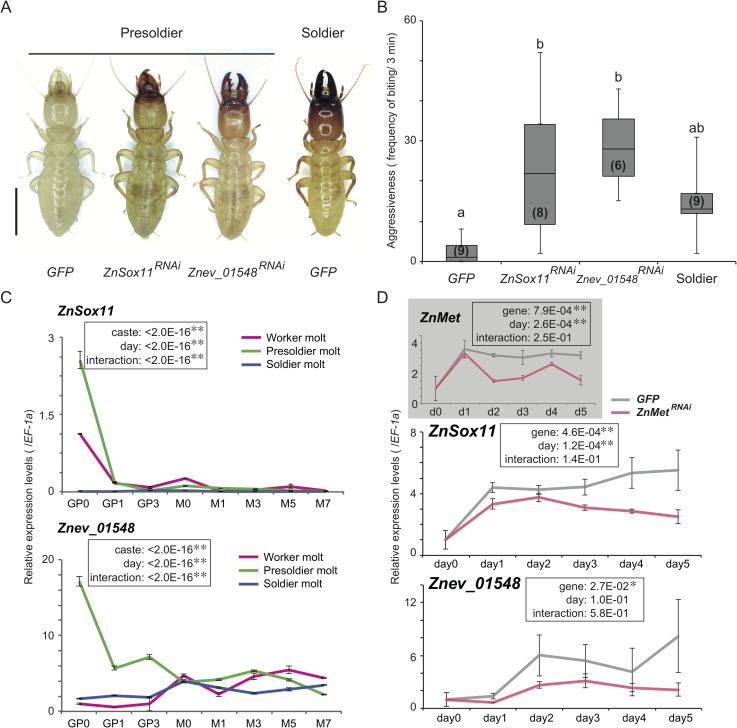
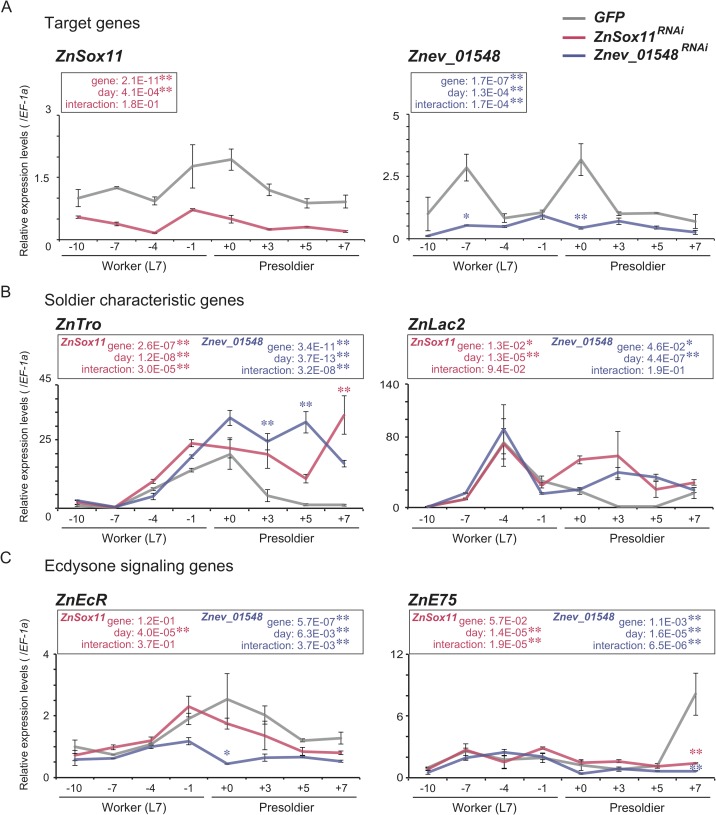
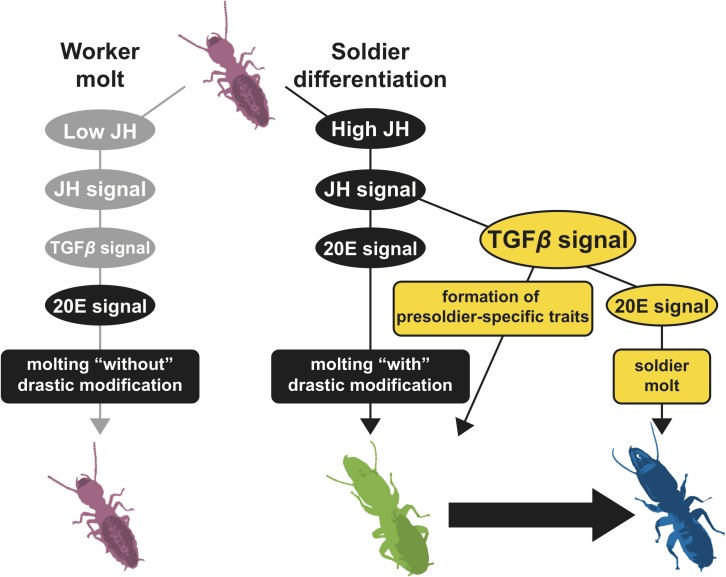
A working knowledge of the proximate factors intrinsic to sterile caste differentiation is necessary to understand the evolution of eusocial insects. Genomic and transcriptomic analyses in social hymenopteran insects have resulted in the hypothesis that sterile castes are generated by the novel function of co-opted or recruited universal gene networks found in solitary ancestors. However, transcriptome analysis during caste differentiation has not been tested in termites, and evolutionary processes associated with acquiring the caste are still unknown. Termites possess the soldier caste, which is regarded as the first acquired permanently sterile caste in the taxon. In this study, we performed a comparative transcriptome analysis in termite heads during 3 molting processes, i.e., worker, presoldier and soldier molts, under natural conditions in an incipient colony of the damp-wood termite Zootermopsis nevadensis. Although similar expression patterns were observed during each molting process, more than 50 genes were shown to be highly expressed before the presoldier (intermediate stage of soldier) molt. We then performed RNA interference (RNAi) of the candidate 13 genes, including transcription factors and uncharacterized protein genes, during presoldier differentiation induced by juvenile hormone (JH) analog treatment. Presoldiers induced after RNAi of two genes related to TGFβ (Transforming growth factor beta) signaling were extremely unusual and possessed soldier-like phenotypes. These individuals also displayed aggressive behaviors similar to natural soldiers when confronted with Formica ants as hypothetical enemies. These presoldiers never molted into the next instar, presumably due to the decreased expression levels of the molting hormone (20-hydroxyecdysone; 20E) signaling genes. These results suggest that TGFβ signaling was acquired for the novel function of regulating between JH and 20E signaling during soldier differentiation in termites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Knockdown of the juvenile hormone receptor gene inhibits soldier-specific morphogenesis in the damp-wood termite Zootermopsis nevadensis (Isoptera: Archotermopsidae).Insect Biochem Mol Biol. 2015 Sep;64:25-31. doi: 10.1016/j.ibmb.2015.07.013. Epub 2015 Jul 15. Insect Biochem Mol Biol. 2015. PMID: 26188329
-
A Crucial Caste Regulation Gene Detected by Comparing Termites and Sister Group Cockroaches.Genetics. 2018 Aug;209(4):1225-1234. doi: 10.1534/genetics.118.301038. Epub 2018 Jun 22. Genetics. 2018. PMID: 29934338 Free PMC article.
-
Histone modifying genes are involved in the molting period during soldier differentiation in Zootermopsis nevadensis.J Insect Physiol. 2019 Aug-Sep;117:103892. doi: 10.1016/j.jinsphys.2019.103892. Epub 2019 Jun 3. J Insect Physiol. 2019. PMID: 31170409
-
The making of the defensive caste: Physiology, development, and evolution of the soldier differentiation in termites.Evol Dev. 2020 Nov;22(6):425-437. doi: 10.1111/ede.12335. Epub 2020 Apr 14. Evol Dev. 2020. PMID: 32291940 Review.
-
Developmental regulation of caste-specific characters in social-insect polyphenism.Evol Dev. 2005 Mar-Apr;7(2):122-9. doi: 10.1111/j.1525-142X.2005.05014.x. Evol Dev. 2005. PMID: 15733310 Review.
Cited by
-
Differential expression of some termite neuropeptides and insulin/IGF-related hormones and their plausible functions in growth, reproduction and caste determination.PeerJ. 2023 Apr 26;11:e15259. doi: 10.7717/peerj.15259. eCollection 2023. PeerJ. 2023. PMID: 37128206 Free PMC article.
-
Diversity of Insect Sesquiterpenoid Regulation.Front Genet. 2020 Sep 10;11:1027. doi: 10.3389/fgene.2020.01027. eCollection 2020. Front Genet. 2020. PMID: 33133135 Free PMC article. Review.
-
Transcriptomic analysis of the honey bee (Apis mellifera) queen spermathecae reveals genes that may be involved in sperm storage after mating.PLoS One. 2021 Jan 8;16(1):e0244648. doi: 10.1371/journal.pone.0244648. eCollection 2021. PLoS One. 2021. PMID: 33417615 Free PMC article.
-
Comparison Between Worker and Soldier Transcriptomes of Termite Neotermes binovatus Reveals Caste Specialization of Host-Flagellate Symbiotic System.Insects. 2025 Mar 19;16(3):325. doi: 10.3390/insects16030325. Insects. 2025. PMID: 40266833 Free PMC article.
-
Impact of Excess Activin A on the Lipids, Metabolites, and Steroids of Adult Mouse Reproductive Organs.Endocrinology. 2025 Mar 24;166(5):bqaf054. doi: 10.1210/endocr/bqaf054. Endocrinology. 2025. PMID: 40209098 Free PMC article.
References
-
- Friedman DA, Gordon DM. Ant Genetics: Reproductive Physiology, Worker Morphology, and Behavior. Ann Rev Neurosci. 2016;39: 41–56. doi: 10.1146/annurev-neuro-070815-013927 - DOI - PubMed
-
- Johnson BR, Jasper WC. Complex patterns of differential expression in candidate master regulatory genes for social behavior in honey bees. Behavioral Ecol Sociobiol. 2016;70: 1033–1043.
-
- Hunt JH, Amdam GV. Bivoltinism as an antecedent to eusociality in the paper wasp genus Polistes. Science. 2005;308: 264–267. doi: 10.1126/science.1109724 - DOI - PMC - PubMed
-
- Hunt JH, Kensinger BJ, Kossuth JA, Henshaw MT, Norberg K, Wolschin F, et al. A diapause pathway underlies the gyne phenotype in Polistes wasps, revealing an evolutionary route to caste-containing insect societies. Proc Natl Acad Sci USA. 2007;104: 14020–14025. doi: 10.1073/pnas.0705660104 - DOI - PMC - PubMed
-
- Jedlička P, Ernst UR, Votavová A, Hanus R, Valterová I. Gene Expression Dynamics in Major Endocrine Regulatory Pathways along the Transition from Solitary to Social Life in a Bumblebee, Bombus terrestris. Front Physiol. 2016;7: 574 doi: 10.3389/fphys.2016.00574 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
