

An m6A-YTH Module Controls Developmental Timing and Morphogenesis in Arabidopsis
- PMID: 29643069
- PMCID: PMC6002192
- DOI: 10.1105/tpc.17.00833
An m6A-YTH Module Controls Developmental Timing and Morphogenesis in Arabidopsis
Abstract
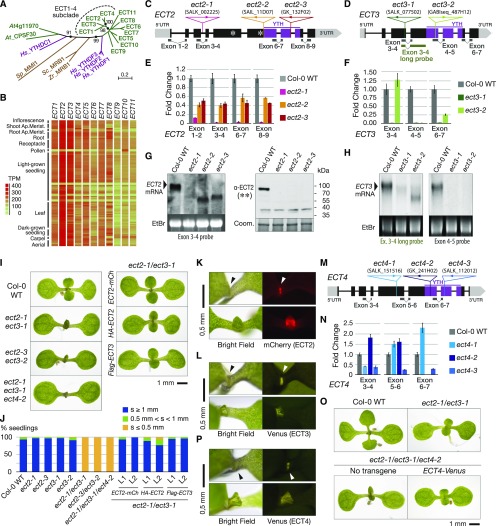
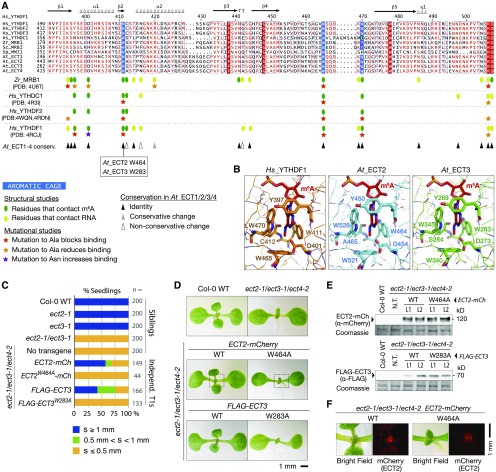
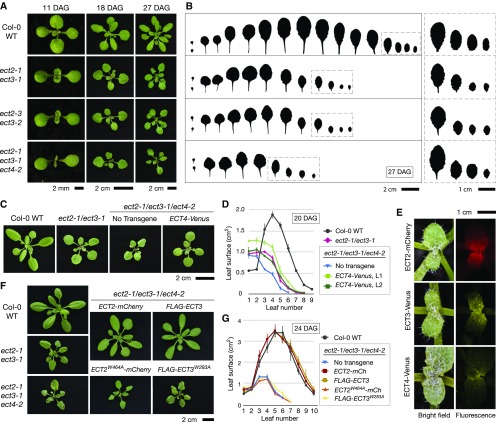
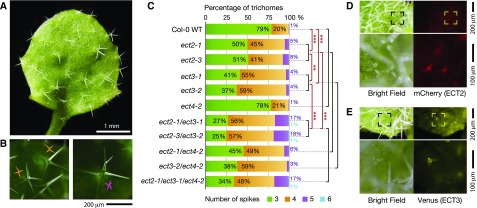
Methylation of N6-adenosine (m6A) in mRNA is an important posttranscriptional gene regulatory mechanism in eukaryotes. m6A provides a binding site for effector proteins ("readers") that influence pre-mRNA splicing, mRNA degradation, or translational efficiency. YT521-B homology (YTH) domain proteins are important m6A readers with established functions in animals. Plants contain more YTH domain proteins than other eukaryotes, but their biological importance remains unknown. Here, we show that the cytoplasmic Arabidopsis thaliana YTH domain proteins EVOLUTIONARILY CONSERVED C-TERMINAL REGION2/3 (ECT2/3) are required for the correct timing of leaf formation and for normal leaf morphology. These functions depend fully on intact m6A binding sites of ECT2 and ECT3, indicating that they function as m6A readers. Mutation of the close ECT2 homolog, ECT4, enhances the delayed leaf emergence and leaf morphology defects of ect2/ect3 mutants, and all three ECT proteins are expressed at leaf formation sites in the shoot apex of young seedlings and in the division zone of developing leaves. ECT2 and ECT3 are also highly expressed at early stages of trichome development and are required for trichome morphology, as previously reported for m6A itself. Overall, our study establishes the relevance of a cytoplasmic m6A-YTH regulatory module in the timing and execution of plant organogenesis.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures

References
-
- Arribas-Hernández L., Marchais A., Poulsen C., Haase B., Hauptmann J., Benes V., Meister G., Brodersen P. (2016). The slicer activity of ARGONAUTE1 is required specifically for the phasing, not production, of trans-acting short interfering RNAs in Arabidopsis. Plant Cell 28: 1563–1580. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
