

Characterizing the Mechanisms of Nonopsonic Uptake of Cryptococci by Macrophages
- PMID: 29643192
- PMCID: PMC5937213
- DOI: 10.4049/jimmunol.1700790
Characterizing the Mechanisms of Nonopsonic Uptake of Cryptococci by Macrophages
Abstract
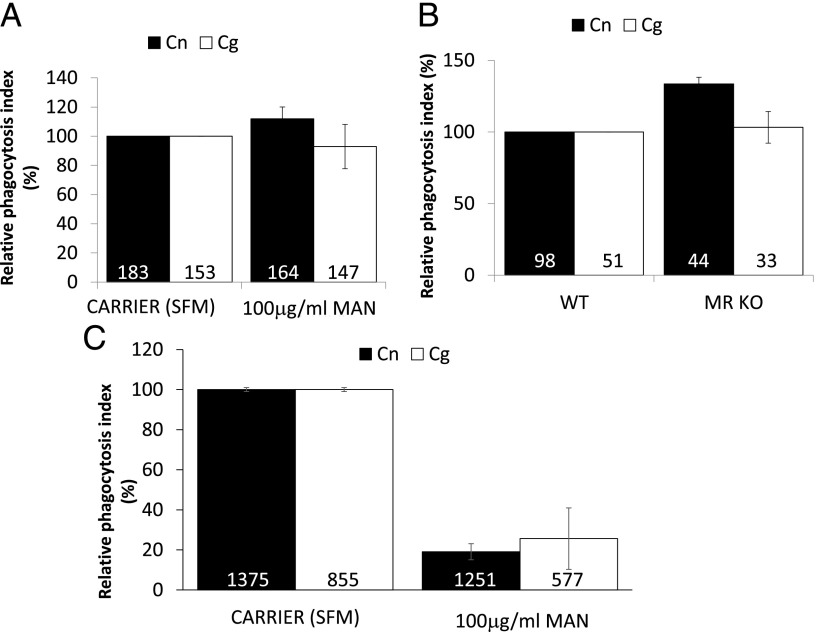
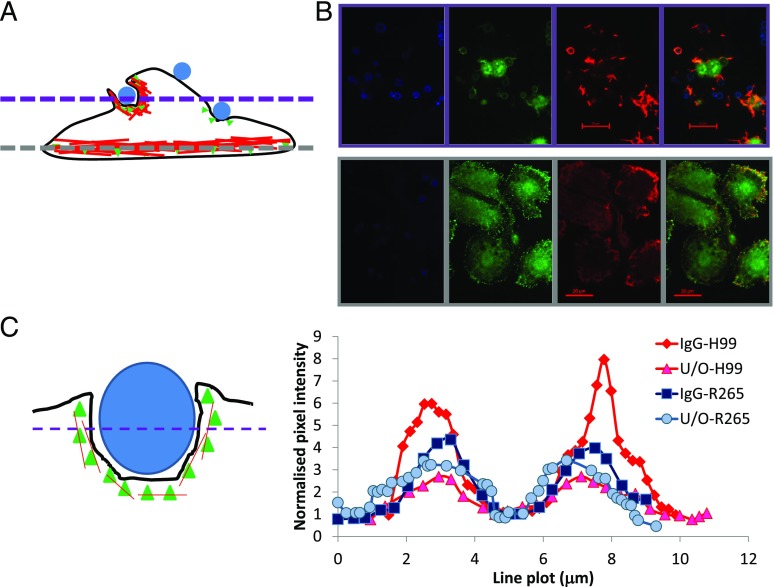
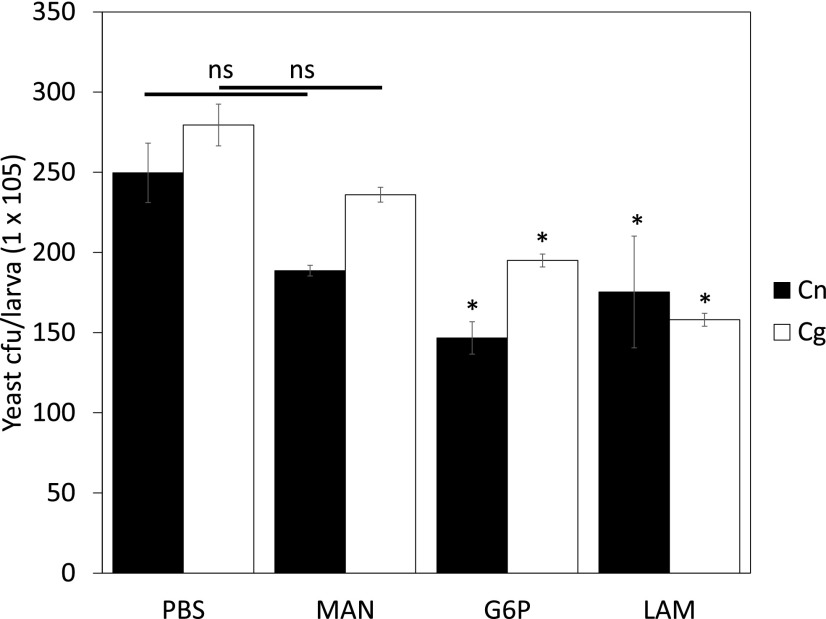
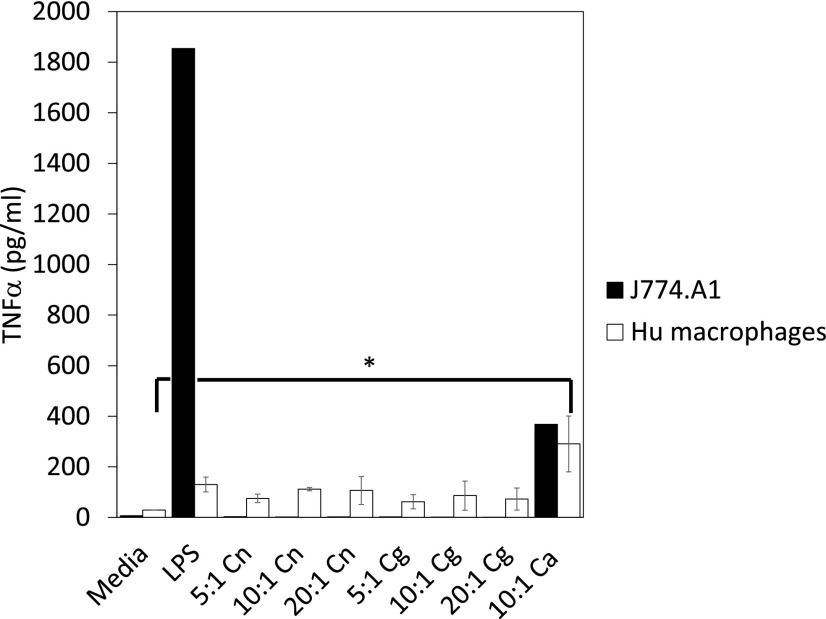
The pathogenic fungus Cryptococcus enters the human host via inhalation into the lung and is able to reside in a niche environment that is serum- (opsonin) limiting. Little is known about the mechanism by which nonopsonic phagocytosis occurs via phagocytes in such situations. Using a combination of soluble inhibitors of phagocytic receptors and macrophages derived from knockout mice and human volunteers, we show that uptake of nonopsonized Cryptococcus neoformans and C. gattii via the mannose receptor is dependent on macrophage activation by cytokines. However, although uptake of C. neoformans is via both dectin-1 and dectin-2, C. gattii uptake occurs largely via dectin-1. Interestingly, dectin inhibitors also blocked phagocytosis of unopsonized Cryptococci in wax moth (Galleria mellonella) larvae and partially protected the larvae from infection by both fungi, supporting a key role for host phagocytes in augmenting early disease establishment. Finally, we demonstrated that internalization of nonopsonized Cryptococci is not accompanied by the nuclear translocation of NF-κB or its concomitant production of proinflammatory cytokines such as TNF-α. Thus, nonopsonized Cryptococci are recognized by mammalian phagocytes in a manner that minimizes proinflammatory cytokine production and potentially facilitates fungal pathogenesis.
Copyright © 2018 The Authors.
Figures




References
-
- Idnurm A., Bahn Y. S., Nielsen K., Lin X., Fraser J. A., Heitman J. 2005. Deciphering the model pathogenic fungus Cryptococcus neoformans. Nat. Rev. Microbiol. 3: 753–764. - PubMed
-
- Feldmesser M., Tucker S., Casadevall A. 2001. Intracellular parasitism of macrophages by Cryptococcus neoformans. Trends Microbiol. 9: 273–278. - PubMed
-
- Flannagan R. S., Jaumouillé V., Grinstein S. 2012. The cell biology of phagocytosis. Annu. Rev. Pathol. 7: 61–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
