

Long-range proton-coupled electron transfer in biological energy conversion: towards mechanistic understanding of respiratory complex I
- PMID: 29643224
- PMCID: PMC5938582
- DOI: 10.1098/rsif.2017.0916
Long-range proton-coupled electron transfer in biological energy conversion: towards mechanistic understanding of respiratory complex I
Abstract
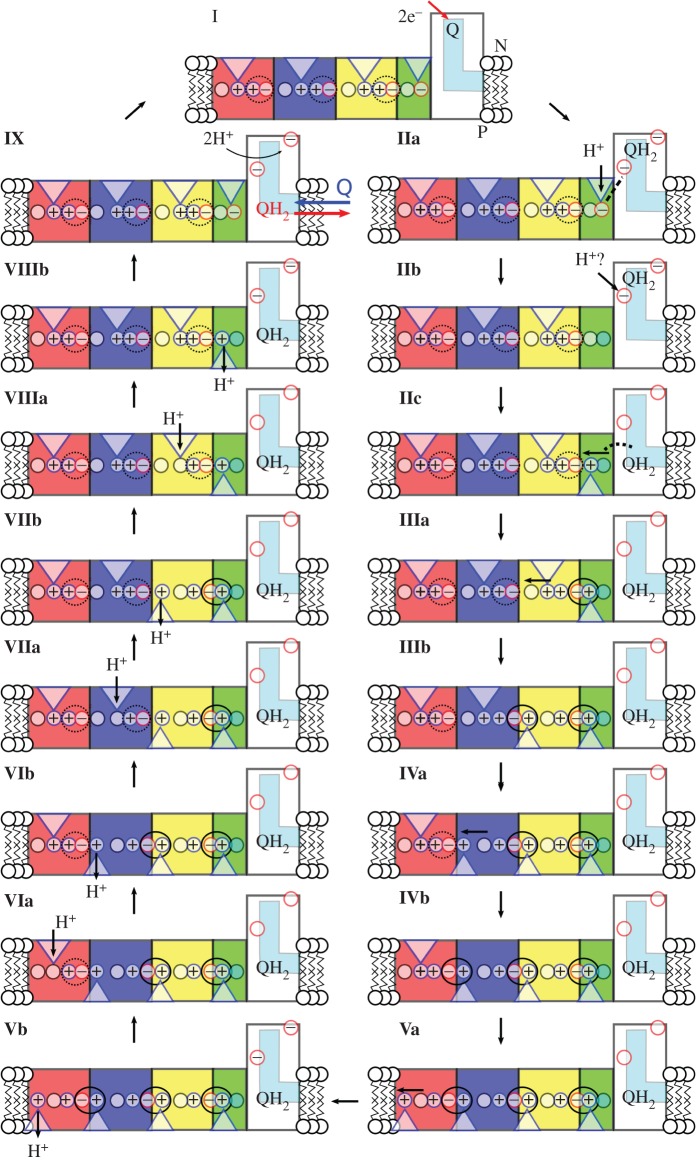
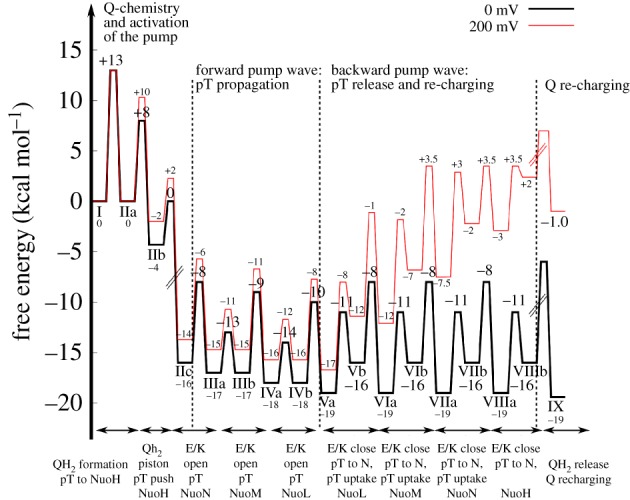
Biological energy conversion is driven by efficient enzymes that capture, store and transfer protons and electrons across large distances. Recent advances in structural biology have provided atomic-scale blueprints of these types of remarkable molecular machinery, which together with biochemical, biophysical and computational experiments allow us to derive detailed energy transduction mechanisms for the first time. Here, I present one of the most intricate and least understood types of biological energy conversion machinery, the respiratory complex I, and how its redox-driven proton-pump catalyses charge transfer across approximately 300 Å distances. After discussing the functional elements of complex I, a putative mechanistic model for its action-at-a-distance effect is presented, and functional parallels are drawn to other redox- and light-driven ion pumps.
Keywords: NADH:ubiquinone oxidoreductase; PCET; bioenergetics; molecular simulations; proton transfer.
© 2018 The Authors.
Conflict of interest statement
I declare I have no competing interests.
Figures
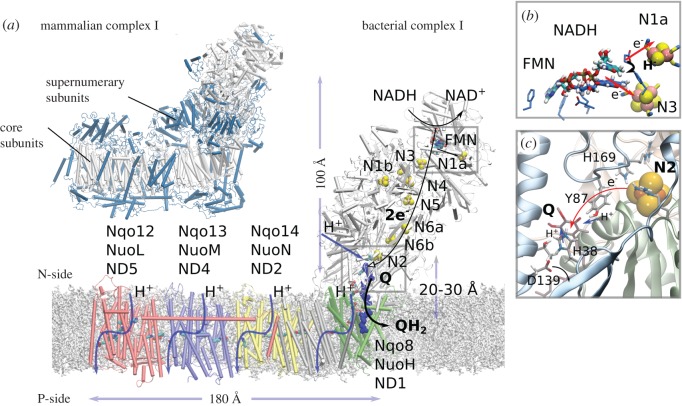
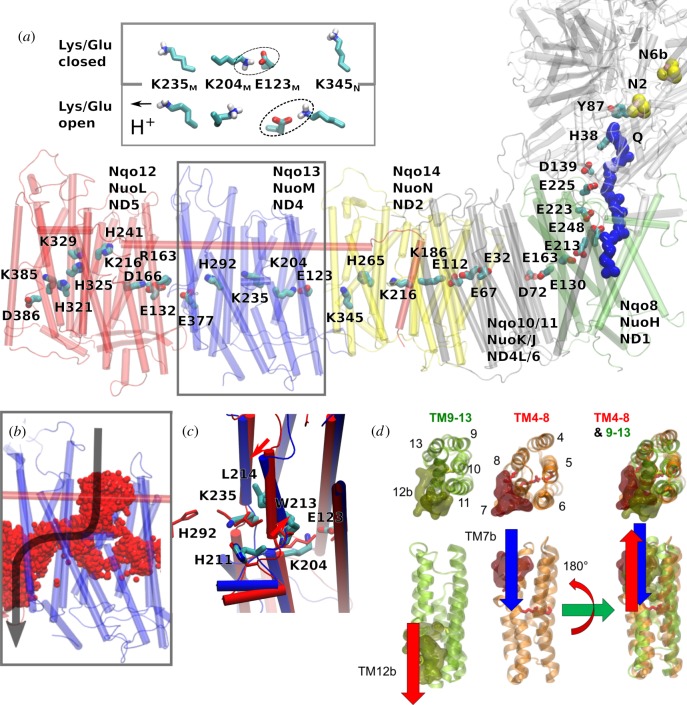
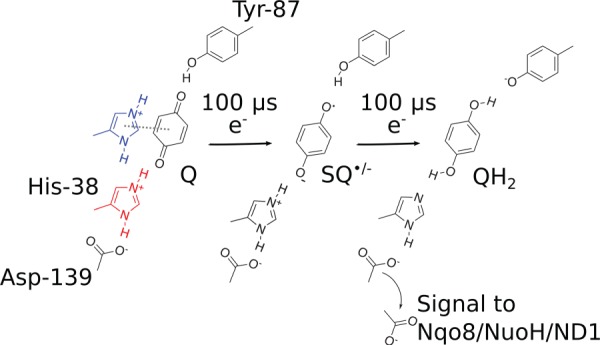
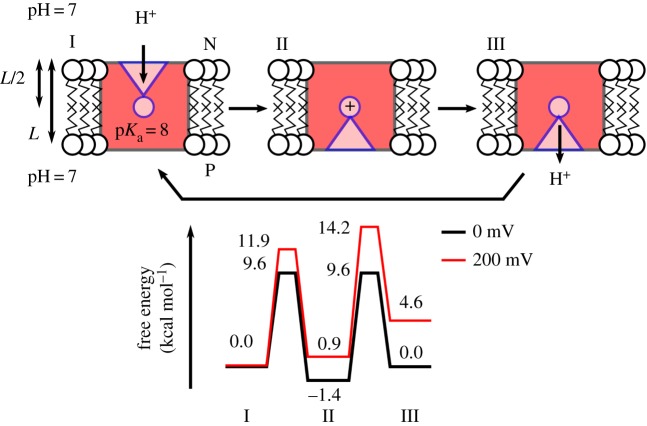
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources