

Identification of a bet-hedging network motif generating noise in hormone concentrations and germination propensity in Arabidopsis
- PMID: 29643226
- PMCID: PMC5938590
- DOI: 10.1098/rsif.2018.0042
Identification of a bet-hedging network motif generating noise in hormone concentrations and germination propensity in Arabidopsis
Abstract
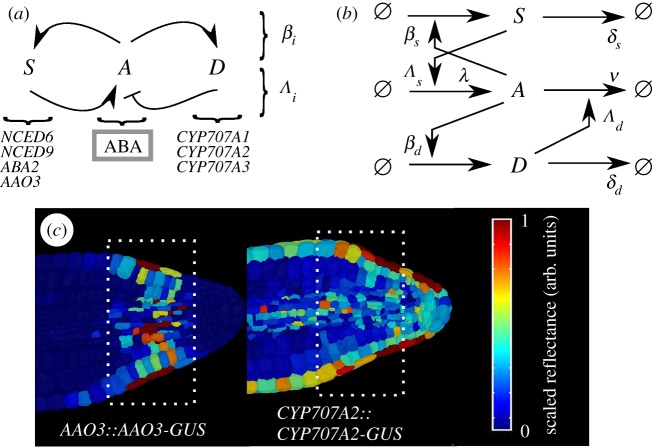
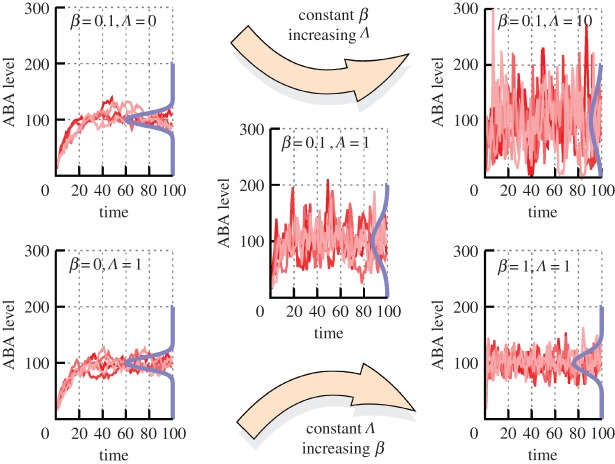
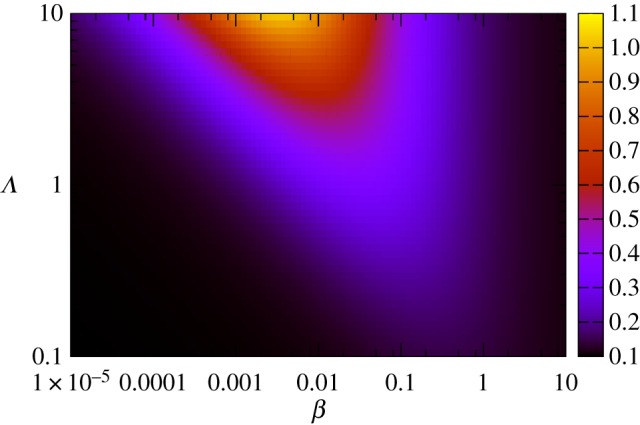
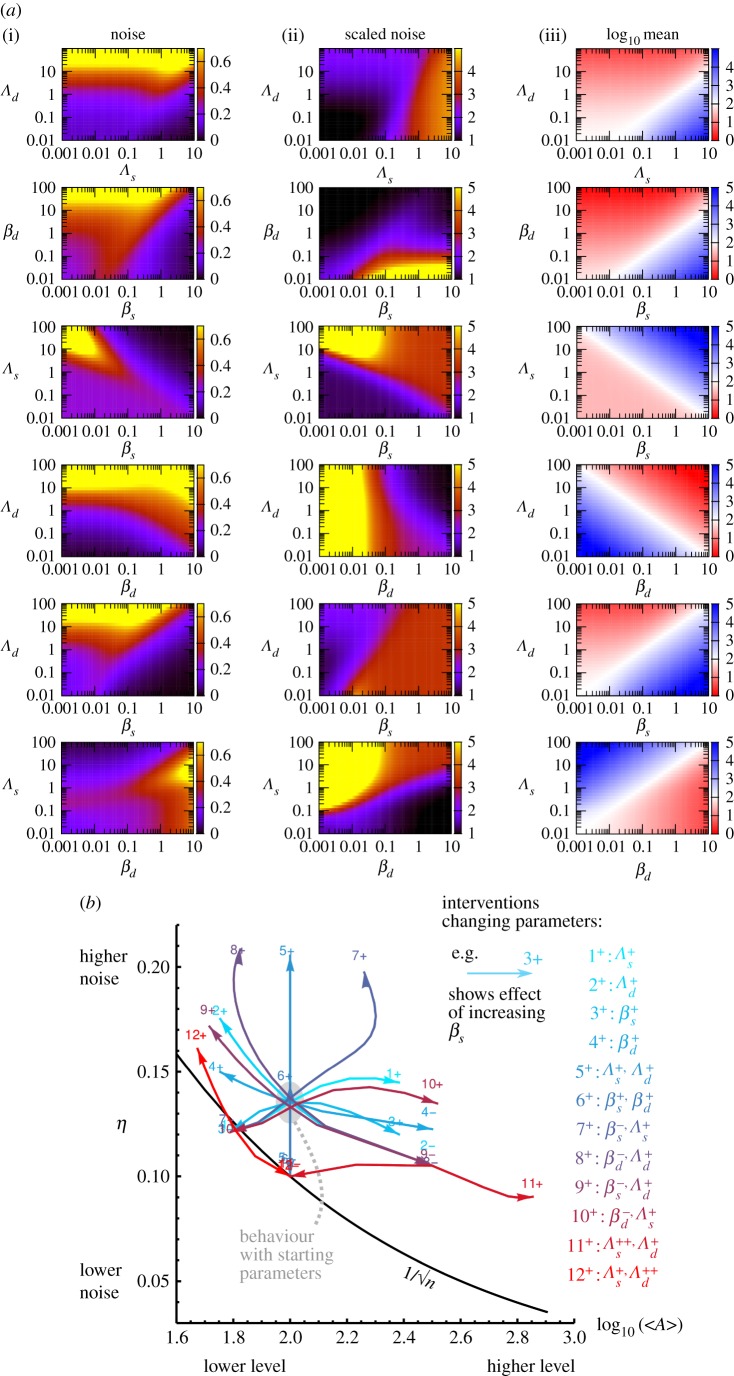
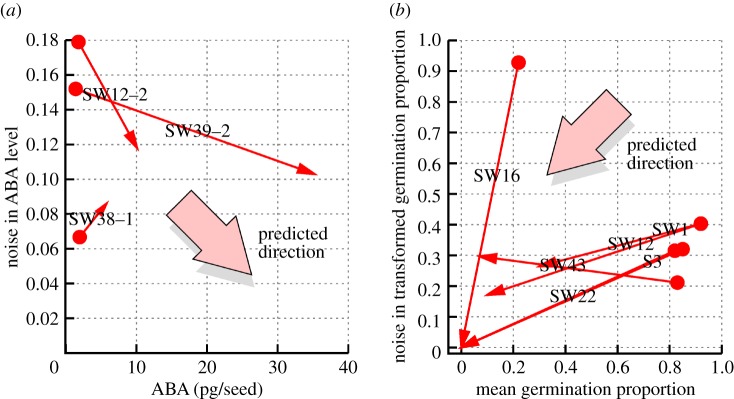
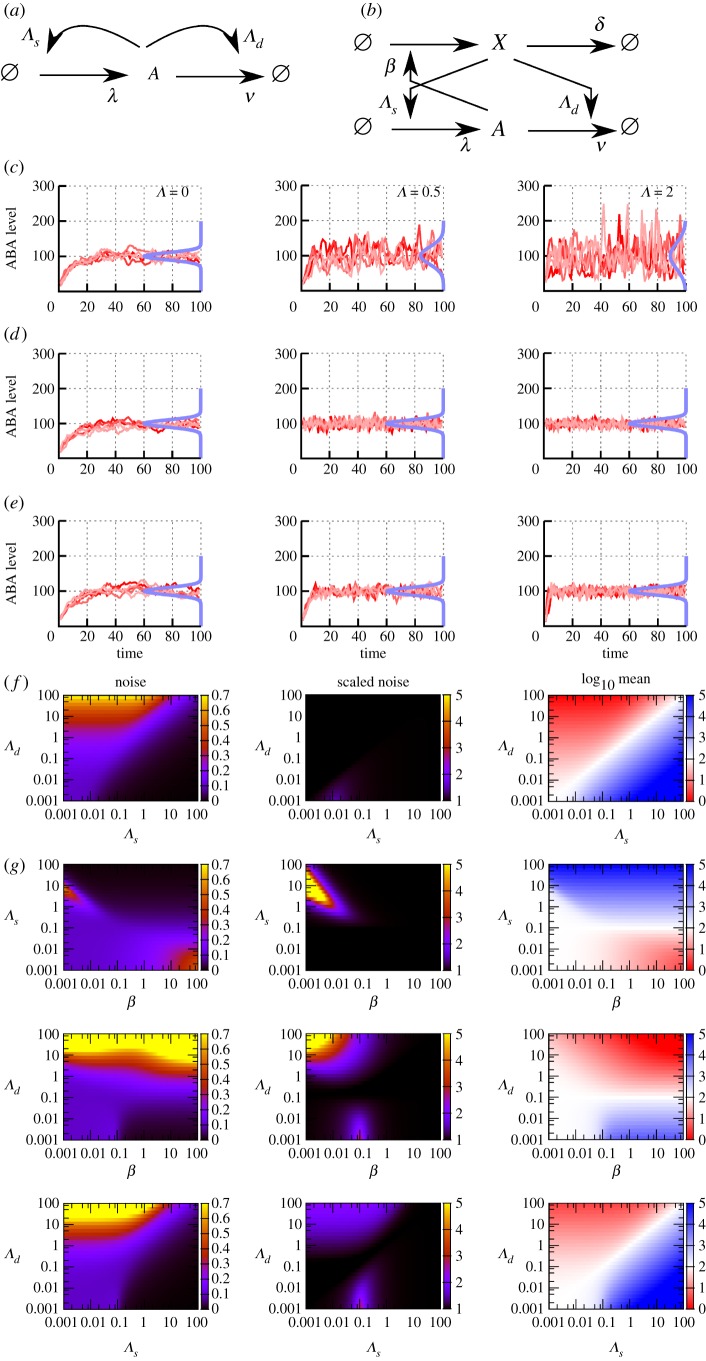
Plants have evolved to exploit stochasticity to hedge bets and ensure robustness to varying environments between generations. In agriculture, environments are more controlled, and this evolved variability decreases potential yields, posing agronomic and food security challenges. Understanding how plant cells generate and harness noise thus presents options for engineering more uniform crop performance. Here, we use stochastic chemical kinetic modelling to analyse a hormone feedback signalling motif in Arabidopsis thaliana seeds that can generate tunable levels of noise in the hormone ABA, governing germination propensity. The key feature of the motif is simultaneous positive feedback regulation of both ABA production and degradation pathways, allowing tunable noise while retaining a constant mean level. We uncover surprisingly rich behaviour underlying the control of levels of, and noise in, ABA abundance. We obtain approximate analytic solutions for steady-state hormone level means and variances under general conditions, showing that antagonistic self-promoting and self-repressing interactions can together be tuned to induce noise while preserving mean hormone levels. We compare different potential architectures for this 'random output generator' with the motif found in Arabidopsis, and report the requirements for tunable control of noise in each case. We identify interventions that may facilitate large decreases in variability in germination propensity, in particular, the turnover of signalling intermediates and the sensitivity of synthesis and degradation machinery, as potentially valuable crop engineering targets.
Keywords: bet-hedging; cell noise; food security; germination; plant hormones; stochastic processes.
© 2018 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures
 .
.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
