

The Functional Oligomeric State of Tegument Protein GP41 Is Essential for Baculovirus Budded Virion and Occlusion-Derived Virion Assembly
- PMID: 29643237
- PMCID: PMC5974493
- DOI: 10.1128/JVI.02083-17
The Functional Oligomeric State of Tegument Protein GP41 Is Essential for Baculovirus Budded Virion and Occlusion-Derived Virion Assembly
Abstract
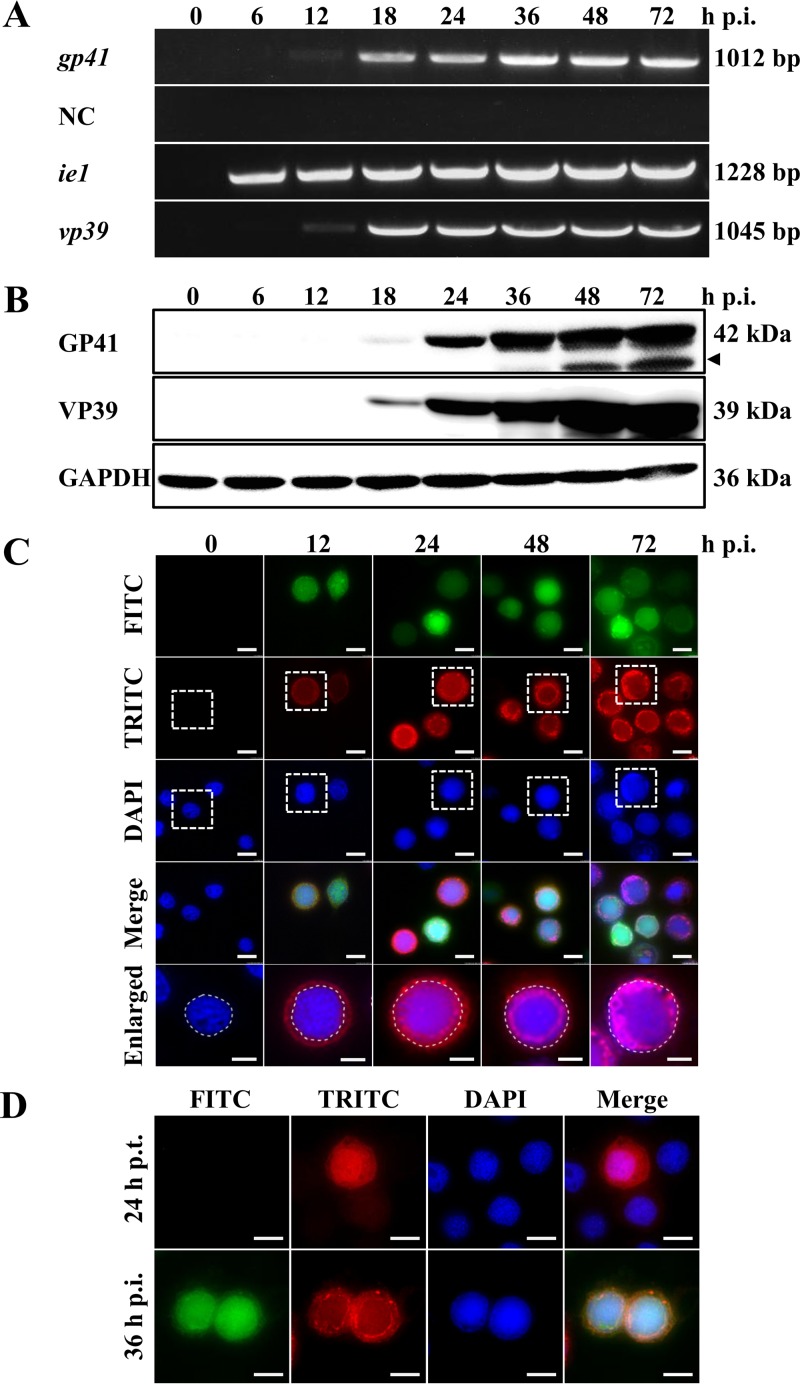
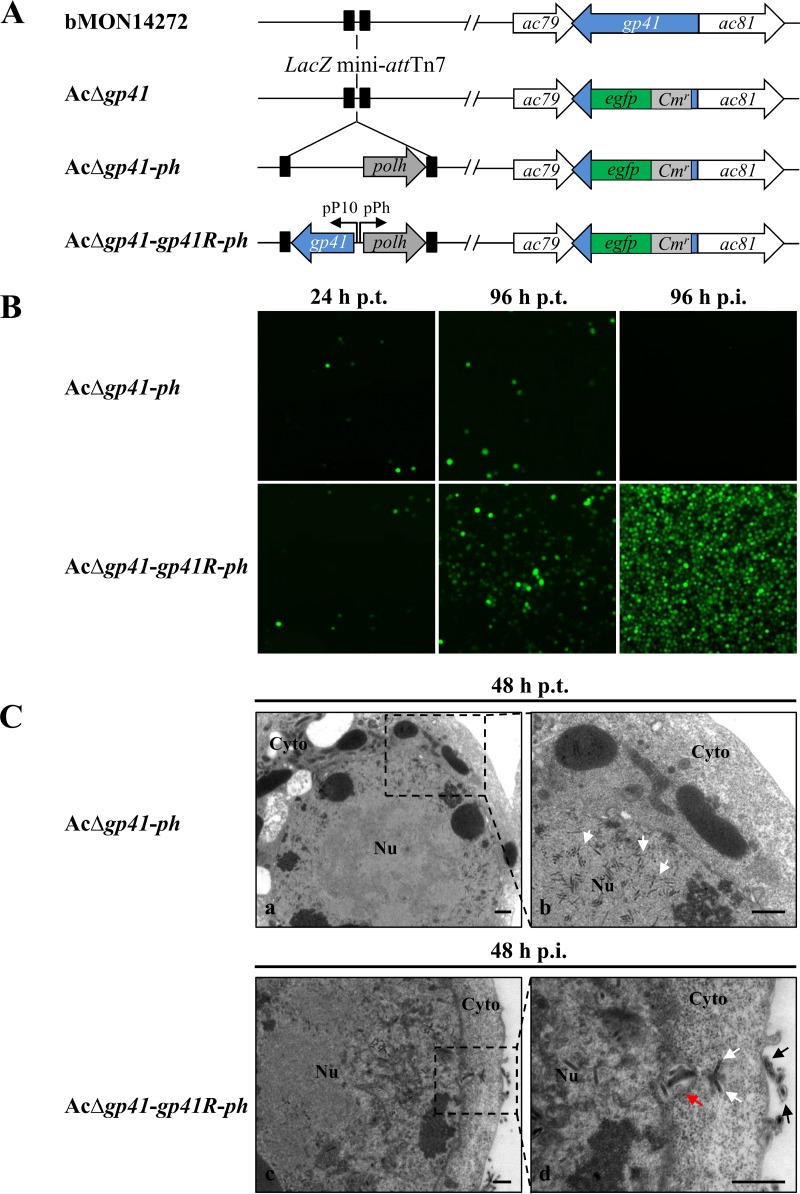
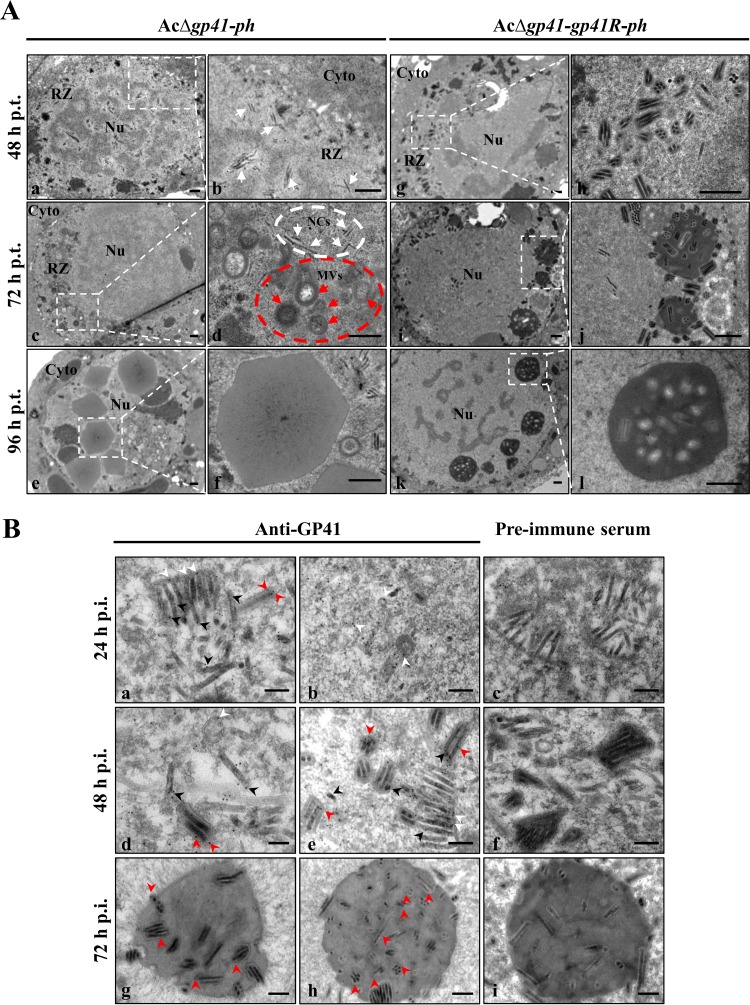
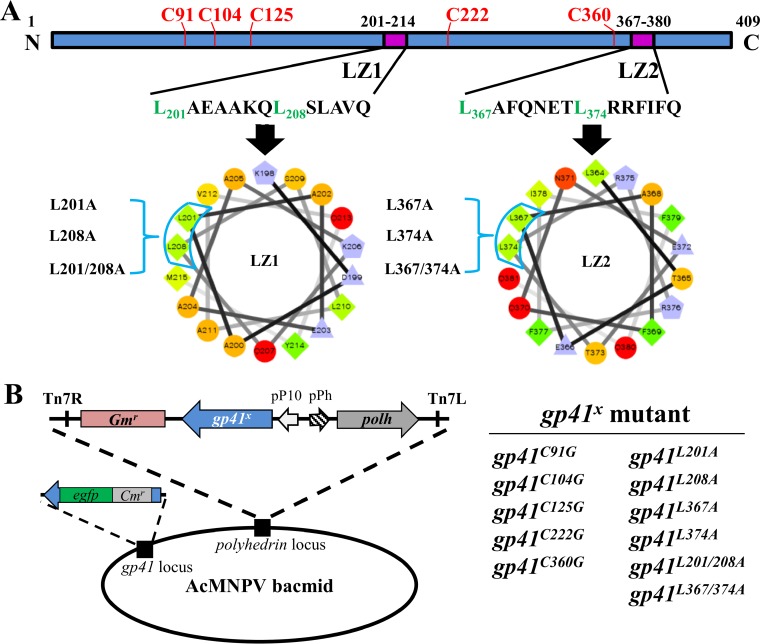
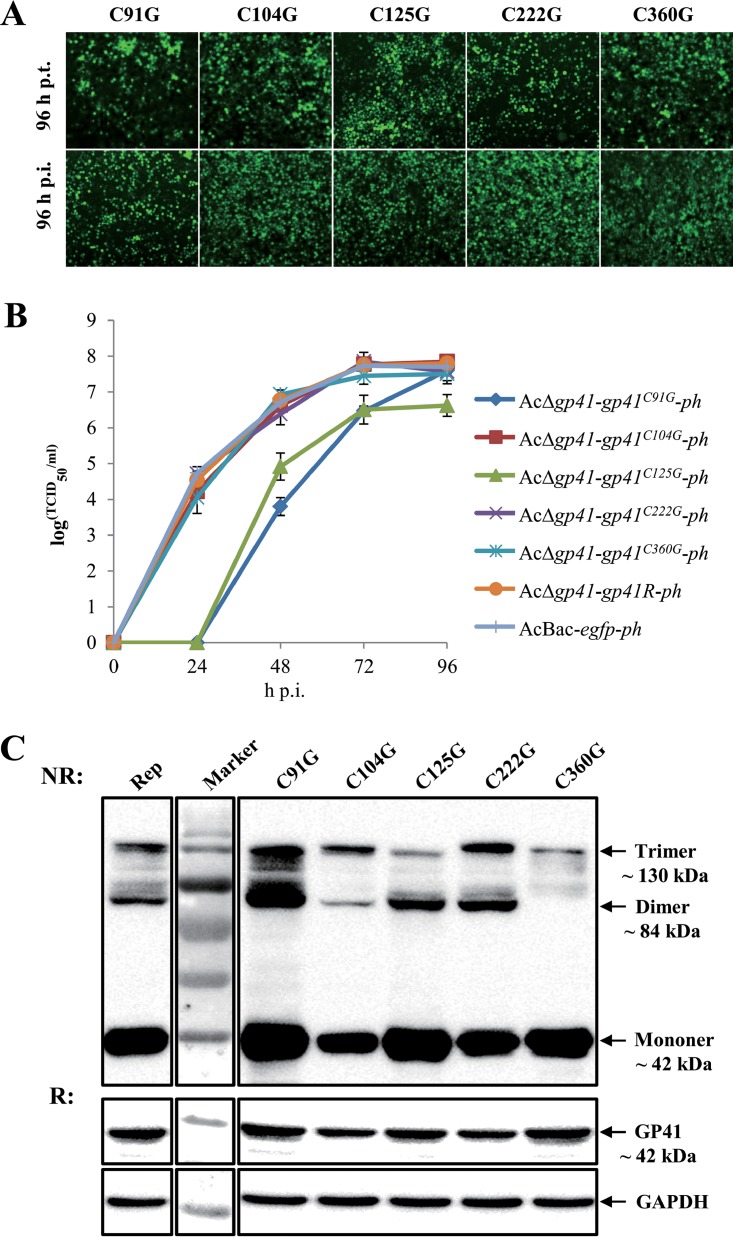
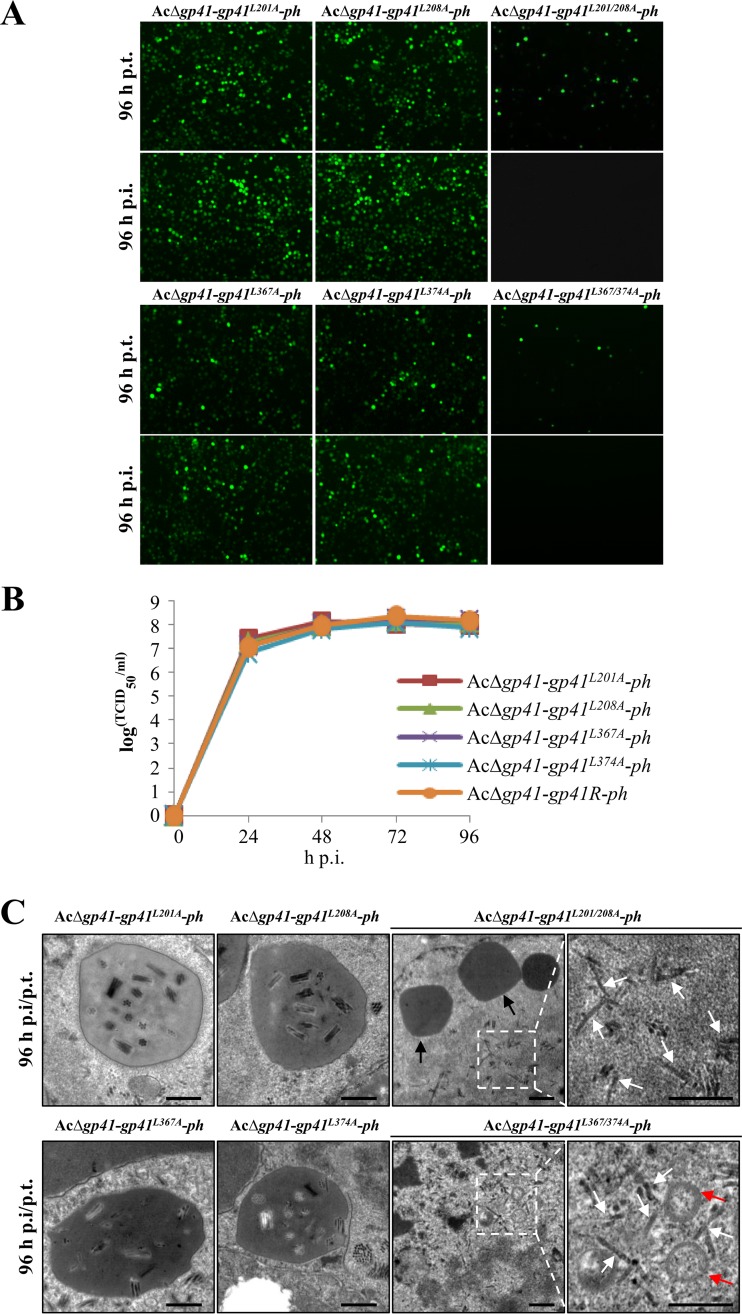
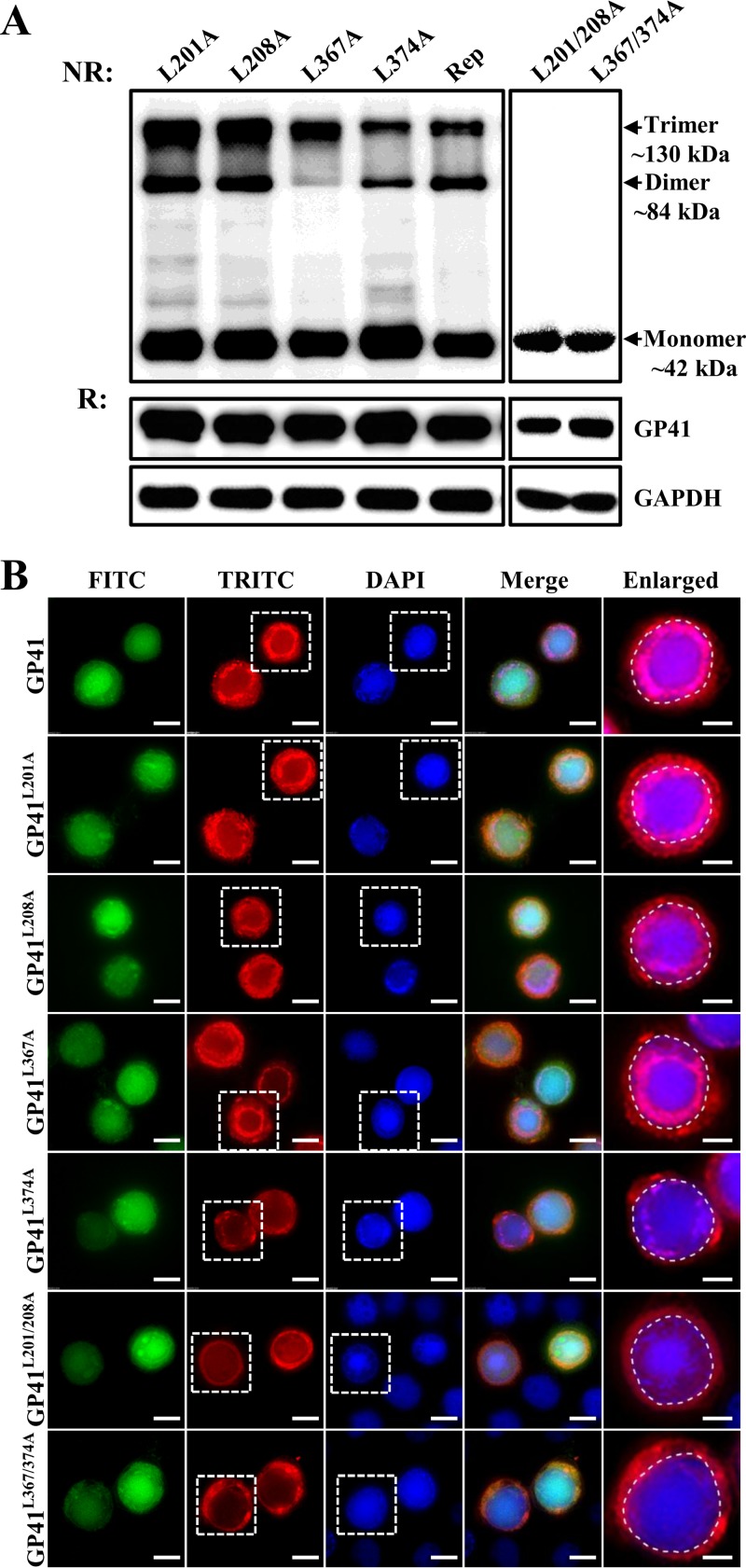
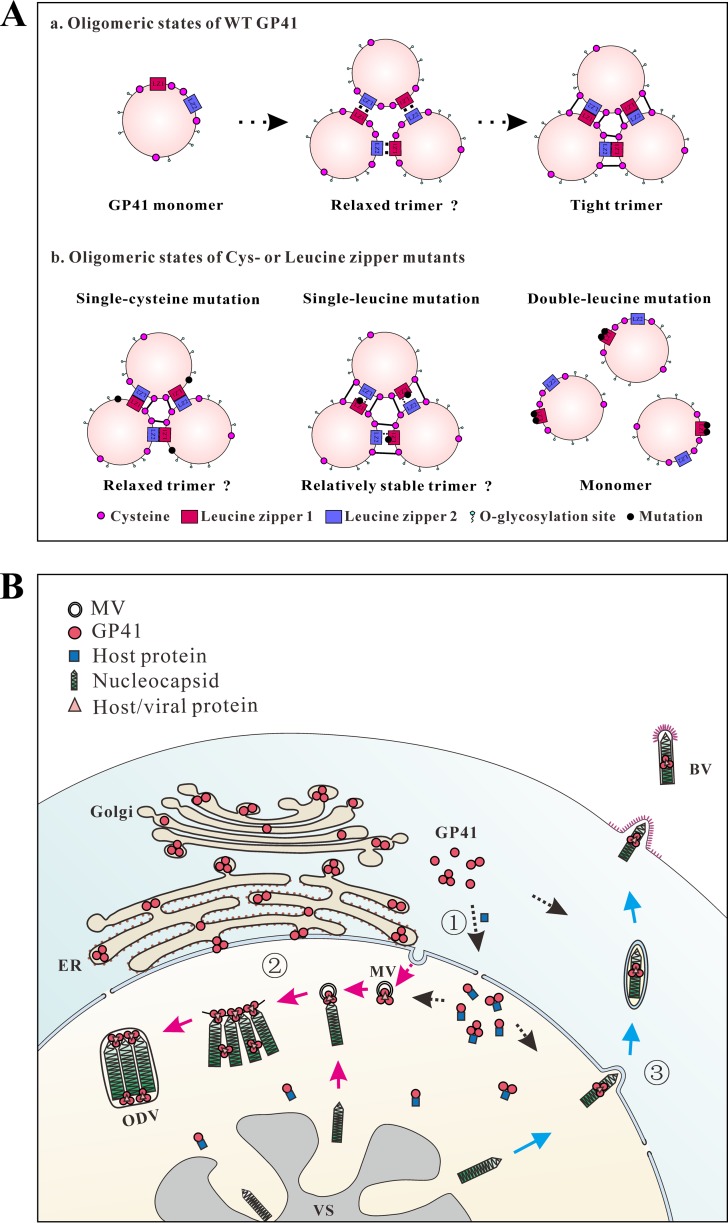
gp41, one of the baculovirus core genes, encodes the only recognized tegument (O-glycosylated) protein of the occlusion-derived virion (ODV) phenotype so far. A previous study using a temperature-sensitive Autographa californica multicapsid nucleopolyhedrovirus (AcMNPV) mutant showed that GP41 plays a crucial role in budded virion (BV) formation. However, the precise function of GP41 in the baculovirus replication cycle remains unclear. In this study, AcMNPV GP41 was found to accumulate around the ring zone (RZ) region within the infected nucleus and finally assembled into both BVs and ODVs. Deletion of gp41 from the AcMNPV genome showed that BVs were no longer formed and ODVs were no longer assembled, suggesting the essential role of this gene in baculovirus virion morphogenesis. In infected cells, besides the 42-kDa monomers, dimers and trimers were detected under nonreducing conditions, whereas only trimeric GP41 forms were selectively incorporated into BVs or ODVs. Mutations of all five cysteines in GP41 individually had minor effects on GP41 oligomer formation, albeit certain mutations impaired infectious BV production, suggesting flexibility in the intermolecular disulfide bonding. Single mutations of key leucines within two predicted leucine zipper-like motifs did not interfere with GP41 oligomerization or BV and ODV formation, but double leucine mutations completely blocked oligomerization of GP41 and progeny BV production. In the latter case, the usual subcellular localization, especially RZ accumulation, of GP41 was abolished. The above findings clearly point out a close correlation between GP41 oligomerization and function and therefore highlight the oligomeric state as the functional form of GP41 in the baculovirus replication cycle.IMPORTANCE The tegument, which is sandwiched between the nucleocapsid and the virion envelope, is an important substructure of many enveloped viruses. It is composed of one or more proteins that have important functions during virus entry, replication, assembly, and egress. Unlike another large DNA virus (herpesvirus) that encodes an extensive set of tegument components, baculoviruses very likely exploit the major tegument protein, GP41, to execute functions in baculovirus virion morphogenesis and assembly. However, the function of this O-glycosylated baculovirus tegument protein remains largely unknown. In this study, we identified trimers as the functional structure of GP41 in baculovirus virion morphogenesis and showed that both disulfide bridging and protein-protein interactions via the two leucine zipper-like domains are involved in the formation of different oligomeric states. This study advances our understanding of the unique viral tegument protein GP41 participating in the life cycle of baculoviruses.
Keywords: GP41; baculovirus; function; leucine zipper; oligomerization; tegument protein; virion assembly.
Copyright © 2018 American Society for Microbiology.
Figures

References
-
- Fiorentini S, Marini E, Caracciolo S, Caruso A. 2006. Functions of the HIV-1 matrix protein p17. New Microbiol 29:1–10. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
