

Structure and regulation of the human INO80-nucleosome complex
- PMID: 29643506
- PMCID: PMC5937682
- DOI: 10.1038/s41586-018-0021-6
Structure and regulation of the human INO80-nucleosome complex
Abstract
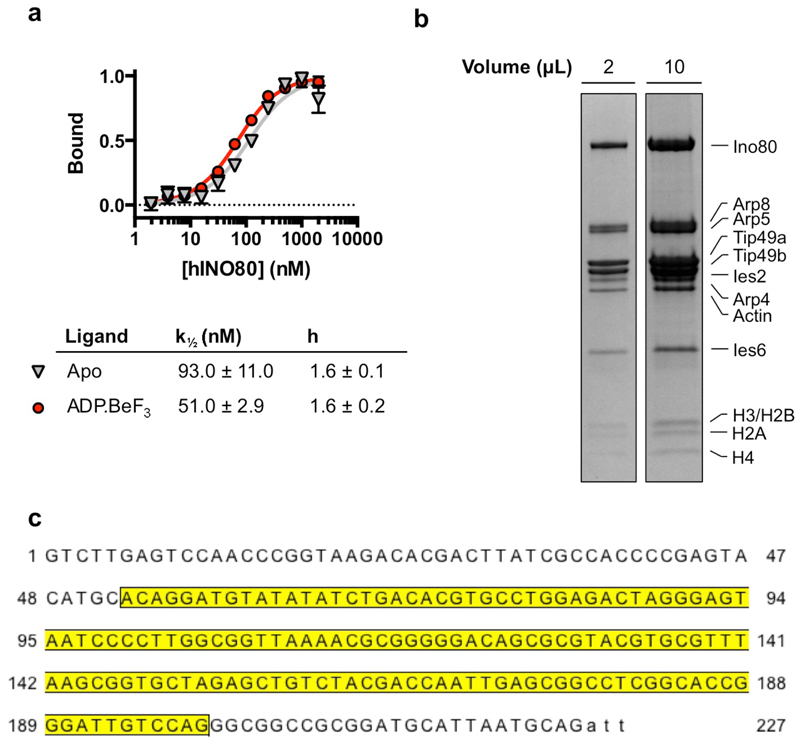
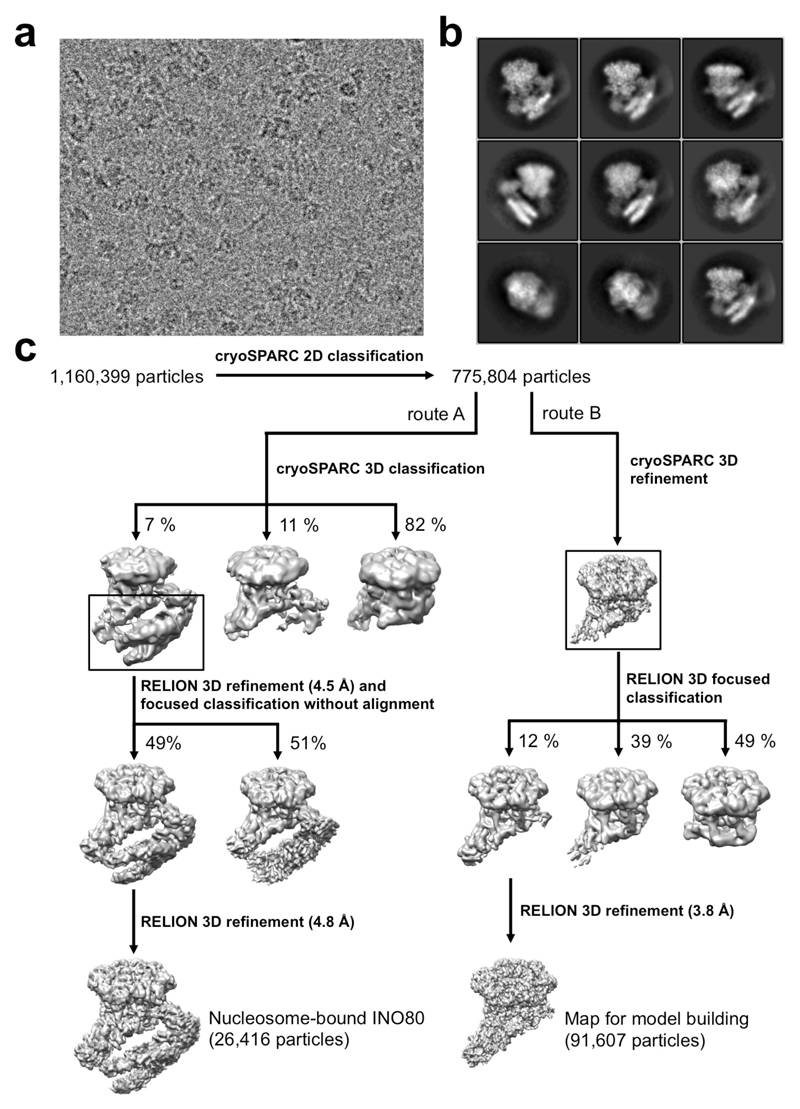
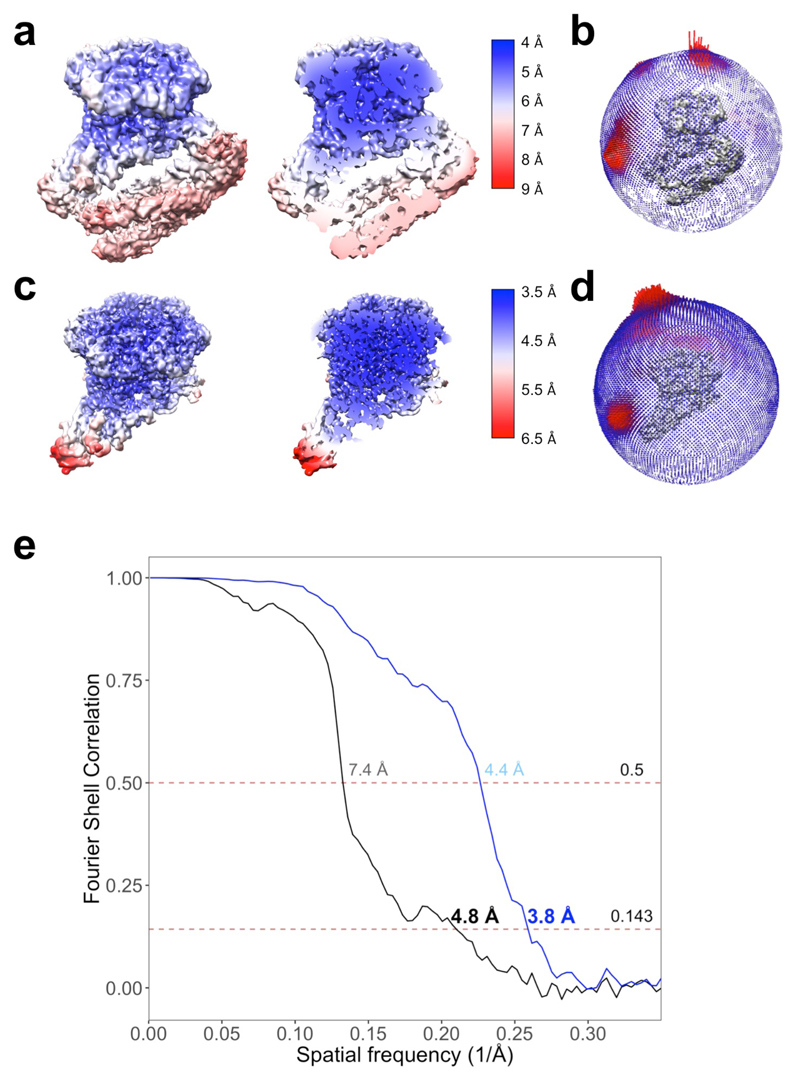
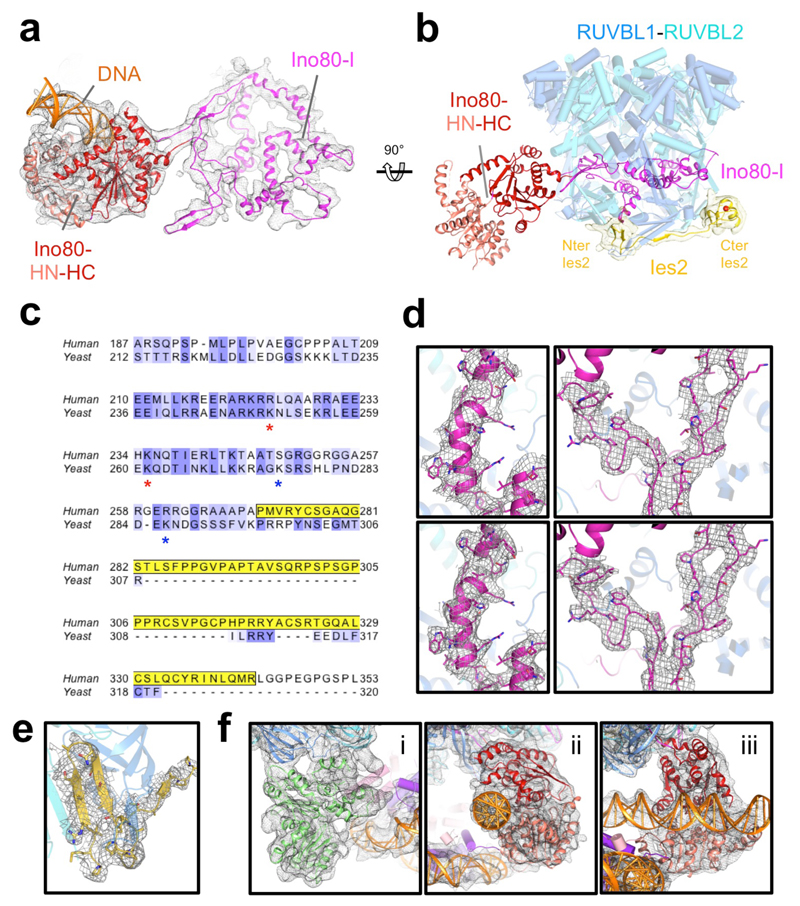
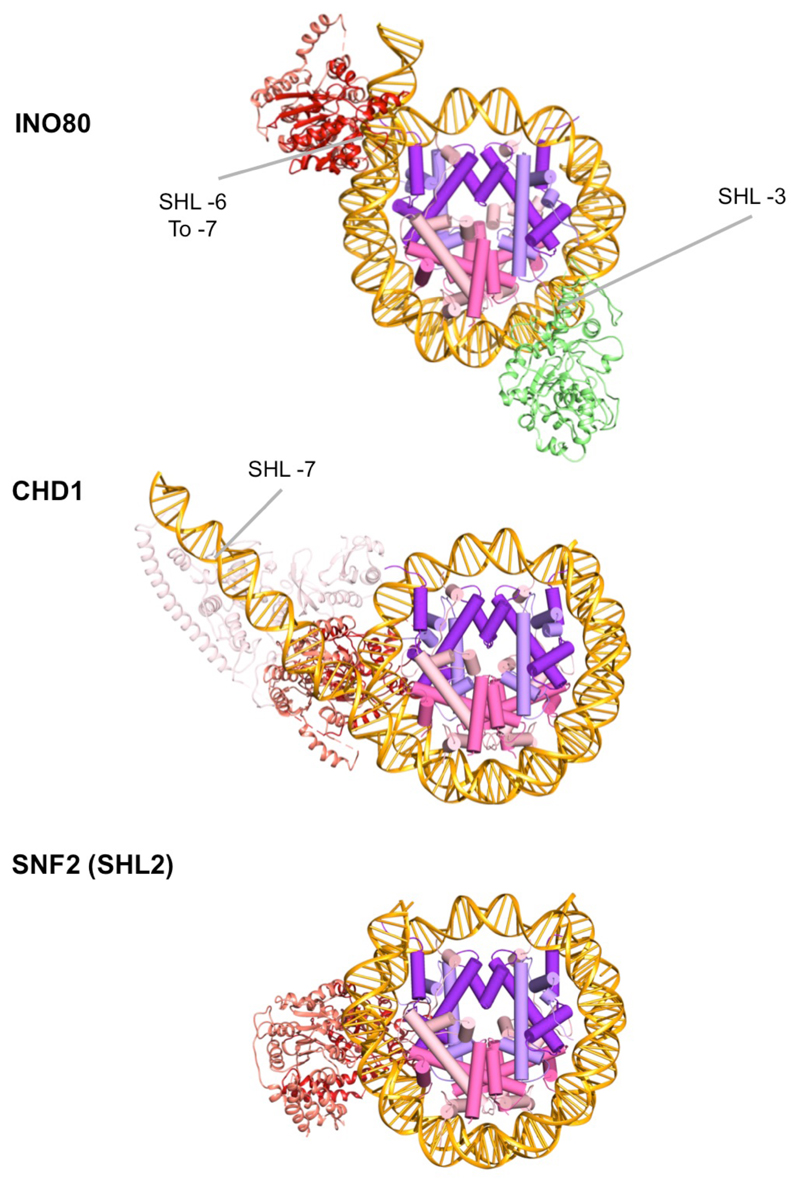
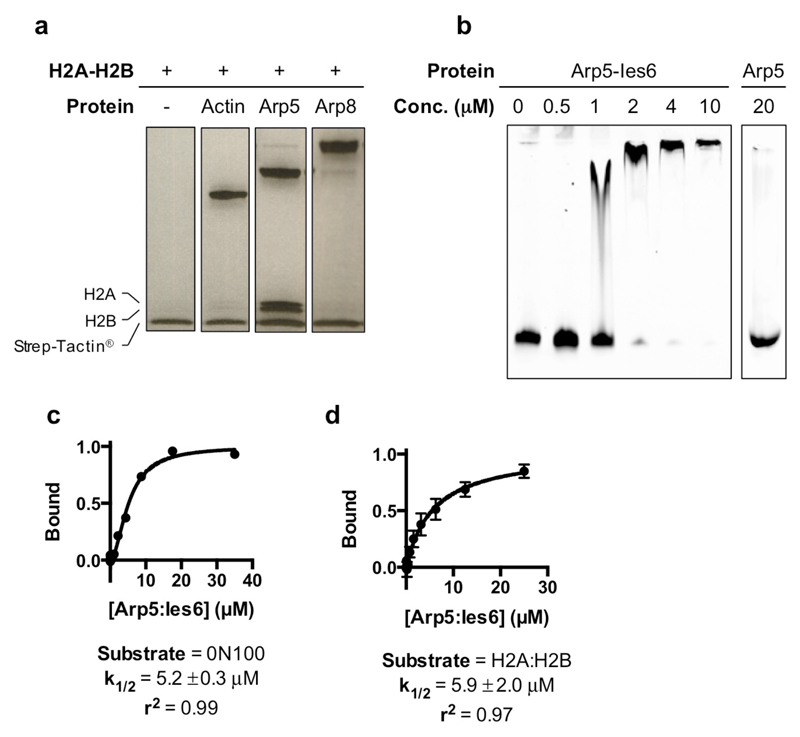
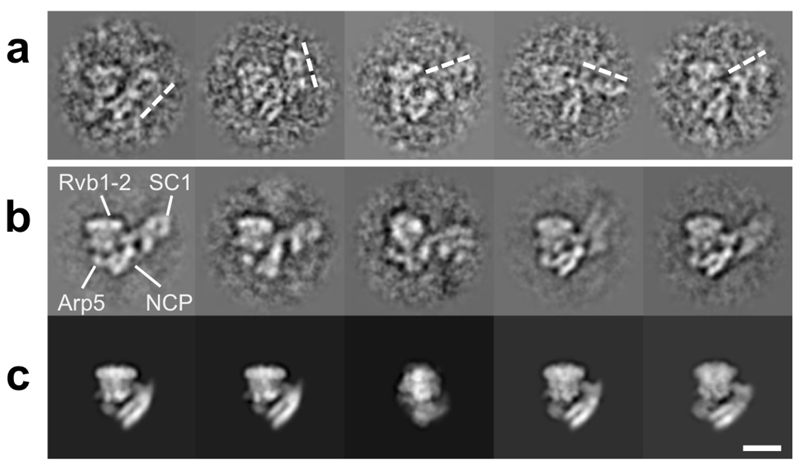
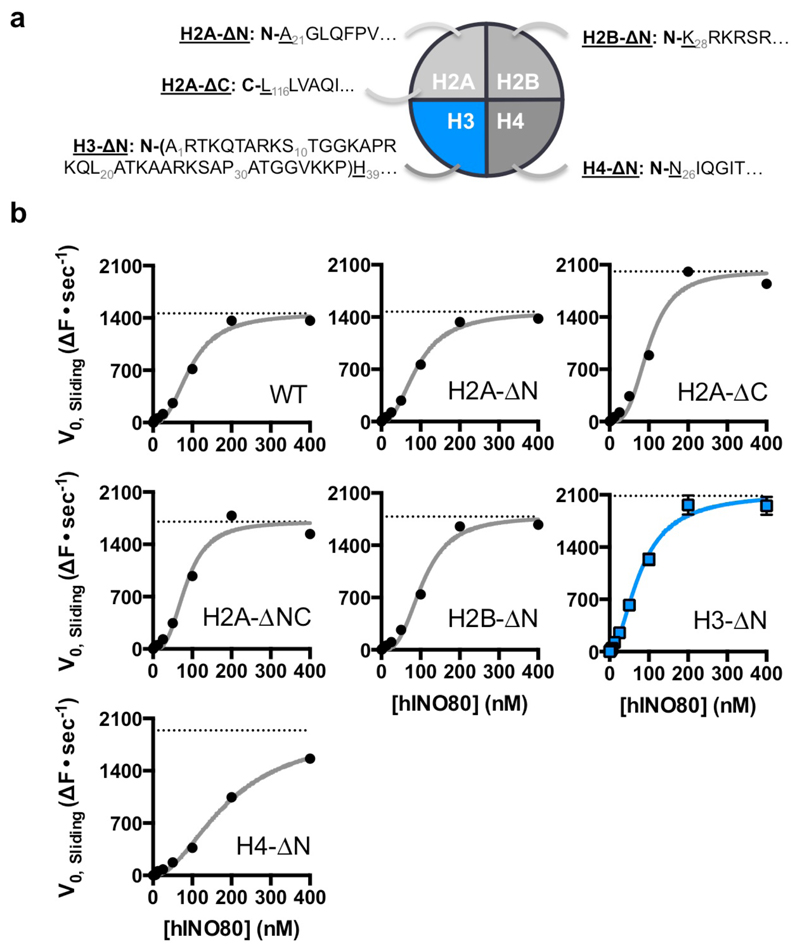
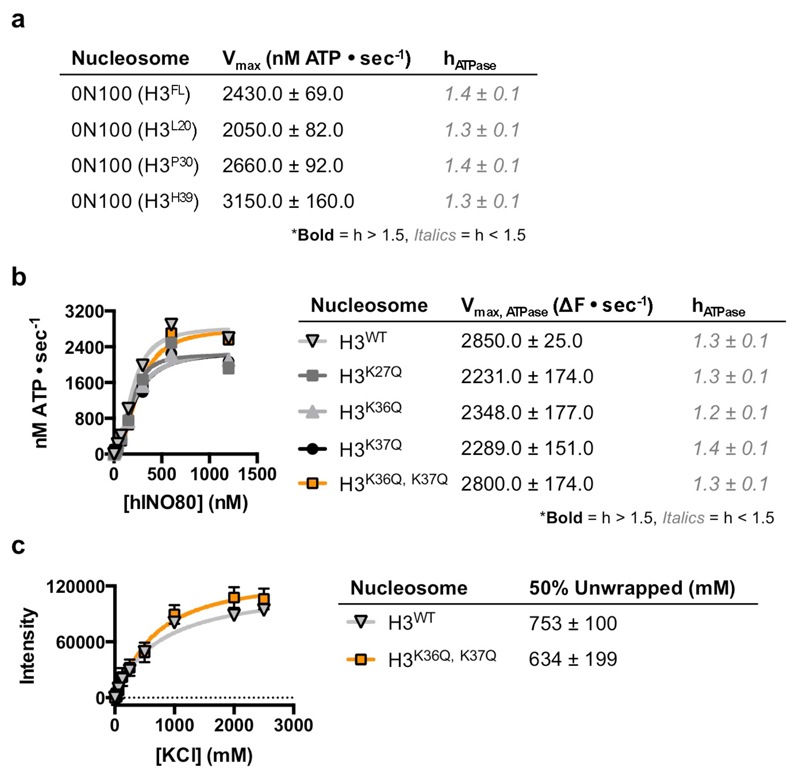
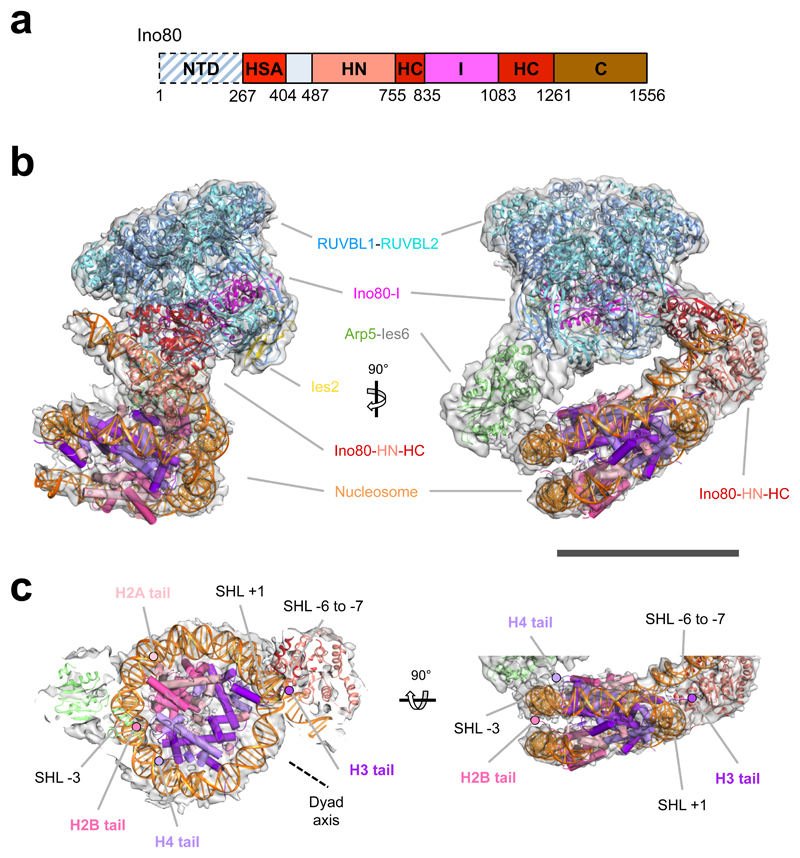
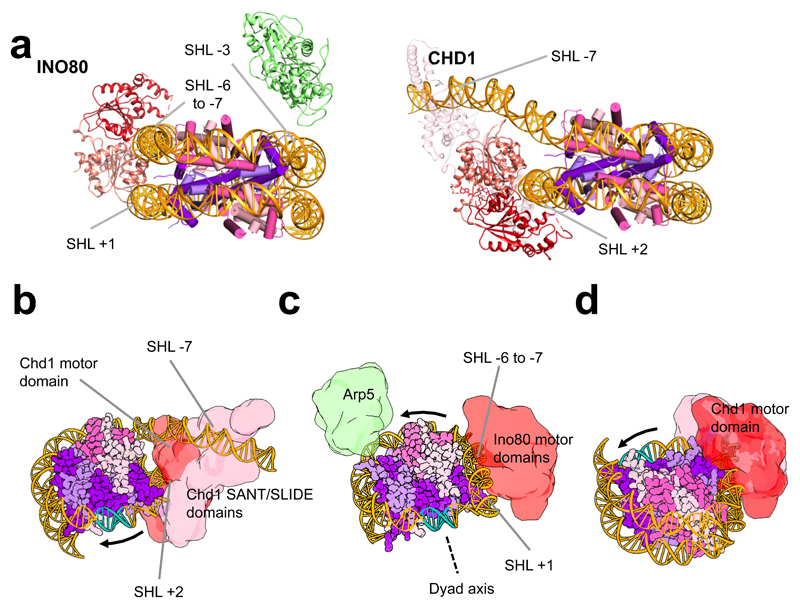
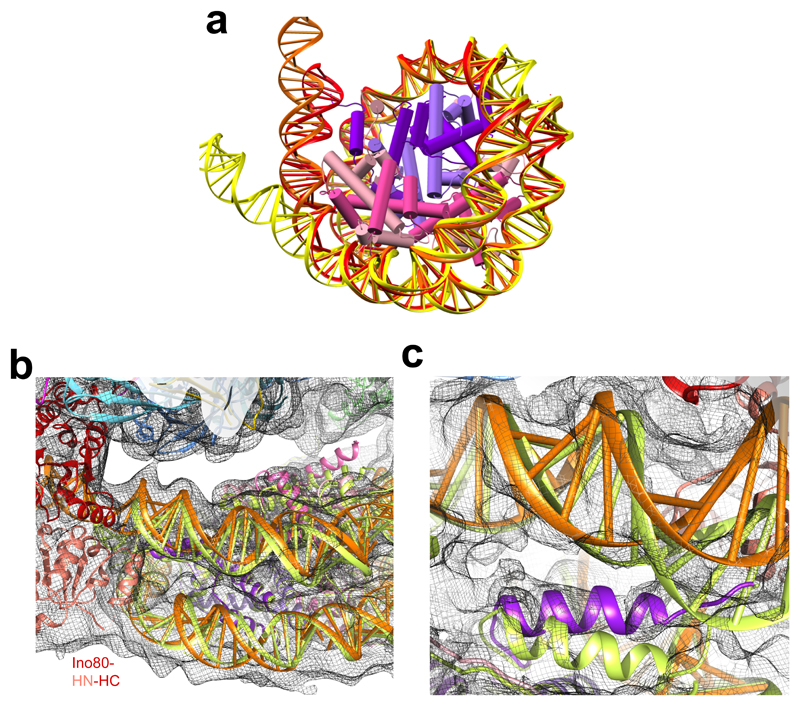
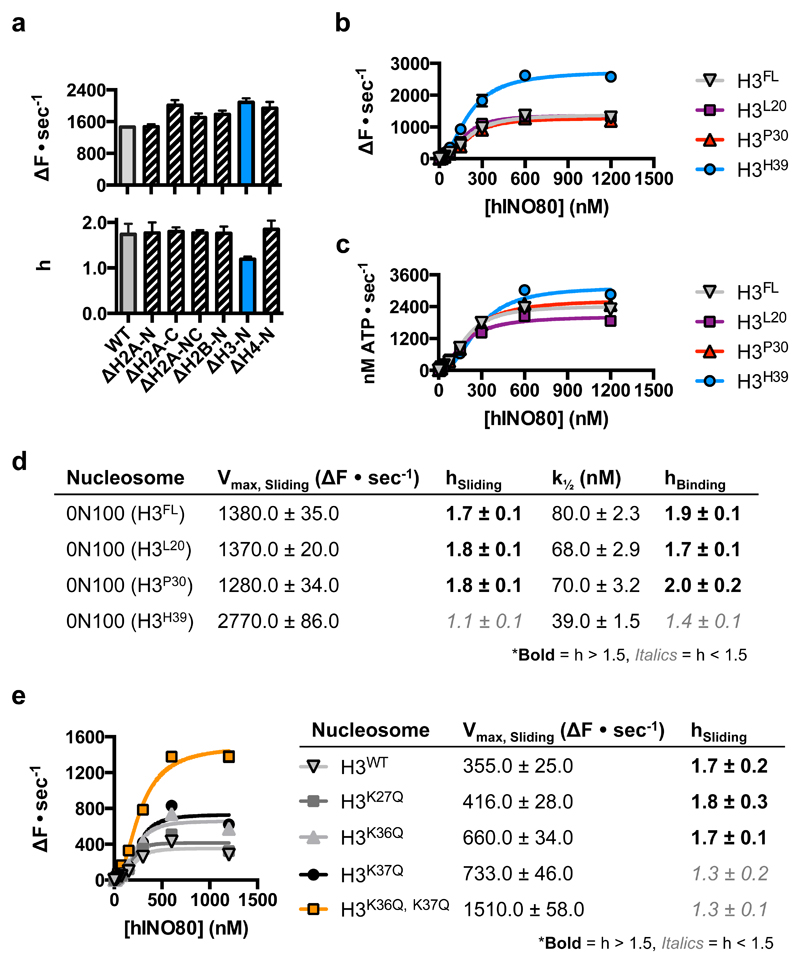
Access to DNA within nucleosomes is required for a variety of processes in cells including transcription, replication and repair. Consequently, cells encode multiple systems that remodel nucleosomes. These complexes can be simple, involving one or a few protein subunits, or more complicated multi-subunit machines 1 . Biochemical studies2-4 have placed the motor domains of several chromatin remodellers in the superhelical location 2 region of the nucleosome. Structural studies of yeast Chd1 and Snf2-a subunit in the complex with the capacity to remodel the structure of chromatin (RSC)-in complex with nucleosomes5-7 have provided insights into the basic mechanism of nucleosome sliding performed by these complexes. However, how larger, multi-subunit remodelling complexes such as INO80 interact with nucleosomes and how remodellers carry out functions such as nucleosome sliding 8 , histone exchange 9 and nucleosome spacing10-12 remain poorly understood. Although some remodellers work as monomers 13 , others work as highly cooperative dimers11, 14, 15. Here we present the structure of the human INO80 chromatin remodeller with a bound nucleosome, which reveals that INO80 interacts with nucleosomes in a previously undescribed manner: the motor domains are located on the DNA at the entry point to the nucleosome, rather than at superhelical location 2. The ARP5-IES6 module of INO80 makes additional contacts on the opposite side of the nucleosome. This arrangement enables the histone H3 tails of the nucleosome to have a role in the regulation of the activities of the INO80 motor domain-unlike in other characterized remodellers, for which H4 tails have been shown to regulate the motor domains.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Structural basis for ATP-dependent chromatin remodelling by the INO80 complex.Nature. 2018 Apr;556(7701):386-390. doi: 10.1038/s41586-018-0029-y. Epub 2018 Apr 11. Nature. 2018. PMID: 29643509 Free PMC article.
-
Crosstalk within a functional INO80 complex dimer regulates nucleosome sliding.Elife. 2017 Jun 6;6:e25782. doi: 10.7554/eLife.25782. Elife. 2017. PMID: 28585918 Free PMC article.
-
INO80 and SWR1 complexes: the non-identical twins of chromatin remodelling.Curr Opin Struct Biol. 2020 Apr;61:50-58. doi: 10.1016/j.sbi.2019.09.002. Epub 2019 Dec 12. Curr Opin Struct Biol. 2020. PMID: 31838293 Free PMC article. Review.
-
Nucleosome-Chd1 structure and implications for chromatin remodelling.Nature. 2017 Oct 26;550(7677):539-542. doi: 10.1038/nature24046. Epub 2017 Oct 11. Nature. 2017. PMID: 29019976 Free PMC article.
-
Energy-driven genome regulation by ATP-dependent chromatin remodellers.Nat Rev Mol Cell Biol. 2024 Apr;25(4):309-332. doi: 10.1038/s41580-023-00683-y. Epub 2023 Dec 11. Nat Rev Mol Cell Biol. 2024. PMID: 38081975 Free PMC article. Review.
Cited by
-
DNA Double Strand Break Repair and Its Control by Nucleosome Remodeling.Front Genet. 2022 Jan 12;12:821543. doi: 10.3389/fgene.2021.821543. eCollection 2021. Front Genet. 2022. PMID: 35096025 Free PMC article. Review.
-
Reorientation of INO80 on hexasomes reveals basis for mechanistic versatility.Science. 2023 Jul 21;381(6655):319-324. doi: 10.1126/science.adf4197. Epub 2023 Jun 29. Science. 2023. PMID: 37384669 Free PMC article.
-
At the Crossroad of Gene Regulation and Genome Organization: Potential Roles for ATP-Dependent Chromatin Remodelers in the Regulation of CTCF-Mediated 3D Architecture.Biology (Basel). 2021 Mar 27;10(4):272. doi: 10.3390/biology10040272. Biology (Basel). 2021. PMID: 33801596 Free PMC article. Review.
-
TBX2 is a neuroblastoma core regulatory circuitry component enhancing MYCN/FOXM1 reactivation of DREAM targets.Nat Commun. 2018 Nov 19;9(1):4866. doi: 10.1038/s41467-018-06699-9. Nat Commun. 2018. PMID: 30451831 Free PMC article.
-
Differential roles of putative arginine fingers of AAA+ ATPases Rvb1 and Rvb2.bioRxiv [Preprint]. 2024 May 13:2024.05.13.593962. doi: 10.1101/2024.05.13.593962. bioRxiv. 2024. PMID: 38798342 Free PMC article. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
