

Innate immune memory in the brain shapes neurological disease hallmarks
- PMID: 29643512
- PMCID: PMC6038912
- DOI: 10.1038/s41586-018-0023-4
Innate immune memory in the brain shapes neurological disease hallmarks
Abstract
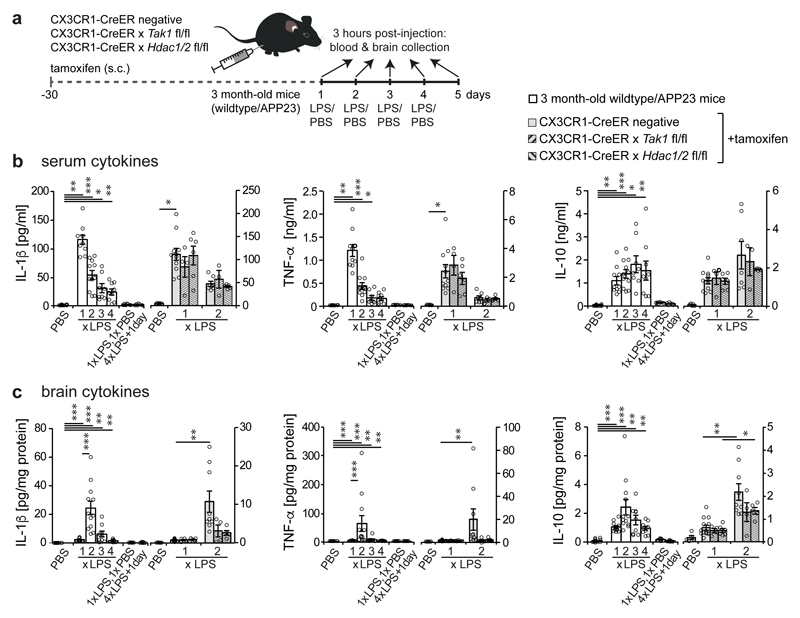
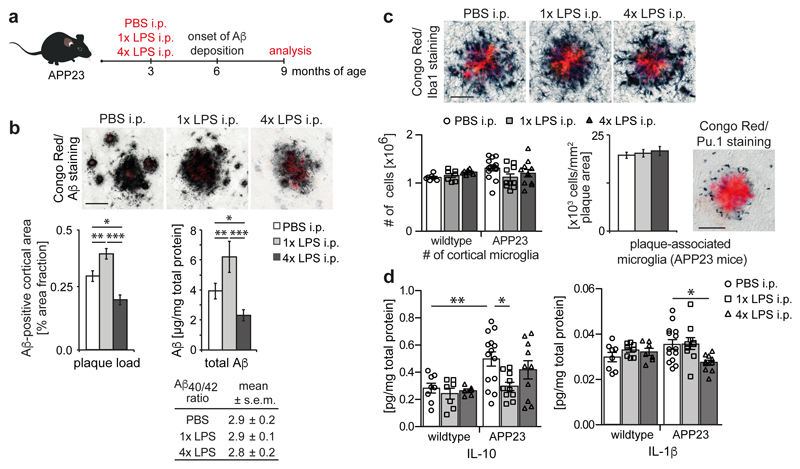
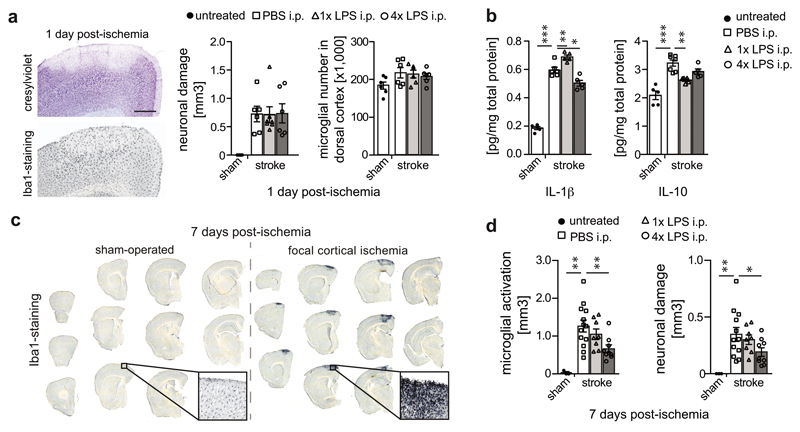
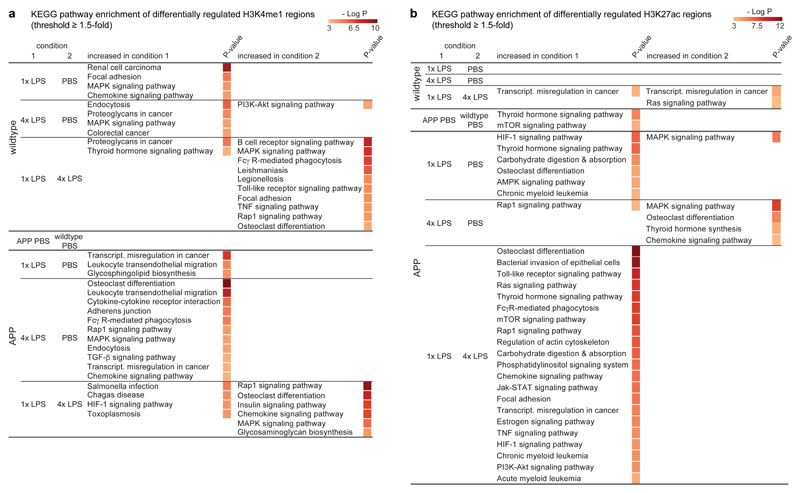
Innate immune memory is a vital mechanism of myeloid cell plasticity that occurs in response to environmental stimuli and alters subsequent immune responses. Two types of immunological imprinting can be distinguished-training and tolerance. These are epigenetically mediated and enhance or suppress subsequent inflammation, respectively. Whether immune memory occurs in tissue-resident macrophages in vivo and how it may affect pathology remains largely unknown. Here we demonstrate that peripherally applied inflammatory stimuli induce acute immune training and tolerance in the brain and lead to differential epigenetic reprogramming of brain-resident macrophages (microglia) that persists for at least six months. Strikingly, in a mouse model of Alzheimer's pathology, immune training exacerbates cerebral β-amyloidosis and immune tolerance alleviates it; similarly, peripheral immune stimulation modifies pathological features after stroke. Our results identify immune memory in the brain as an important modifier of neuropathology.
Conflict of interest statement
The authors have no competing financial interests.
Figures
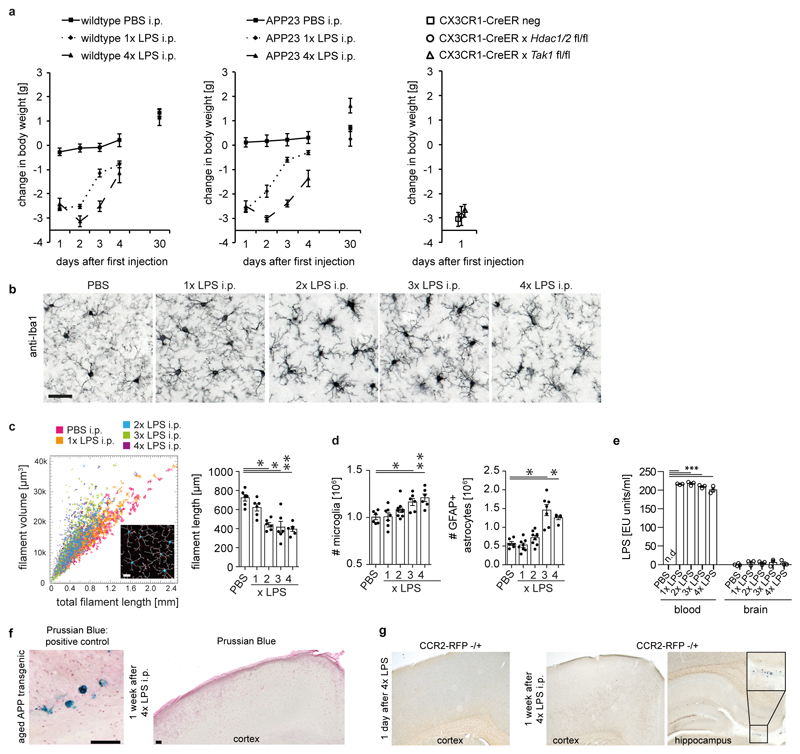

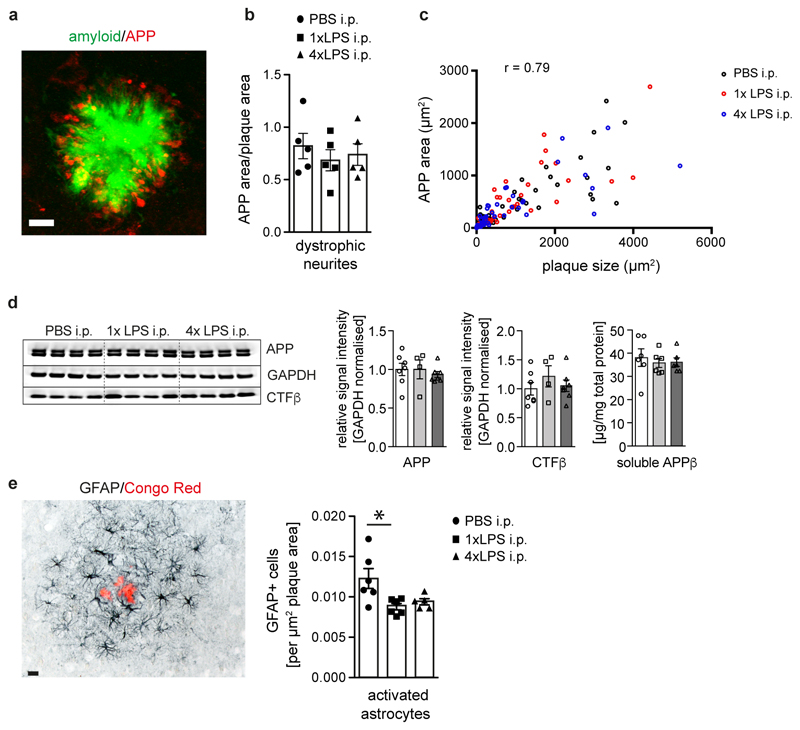
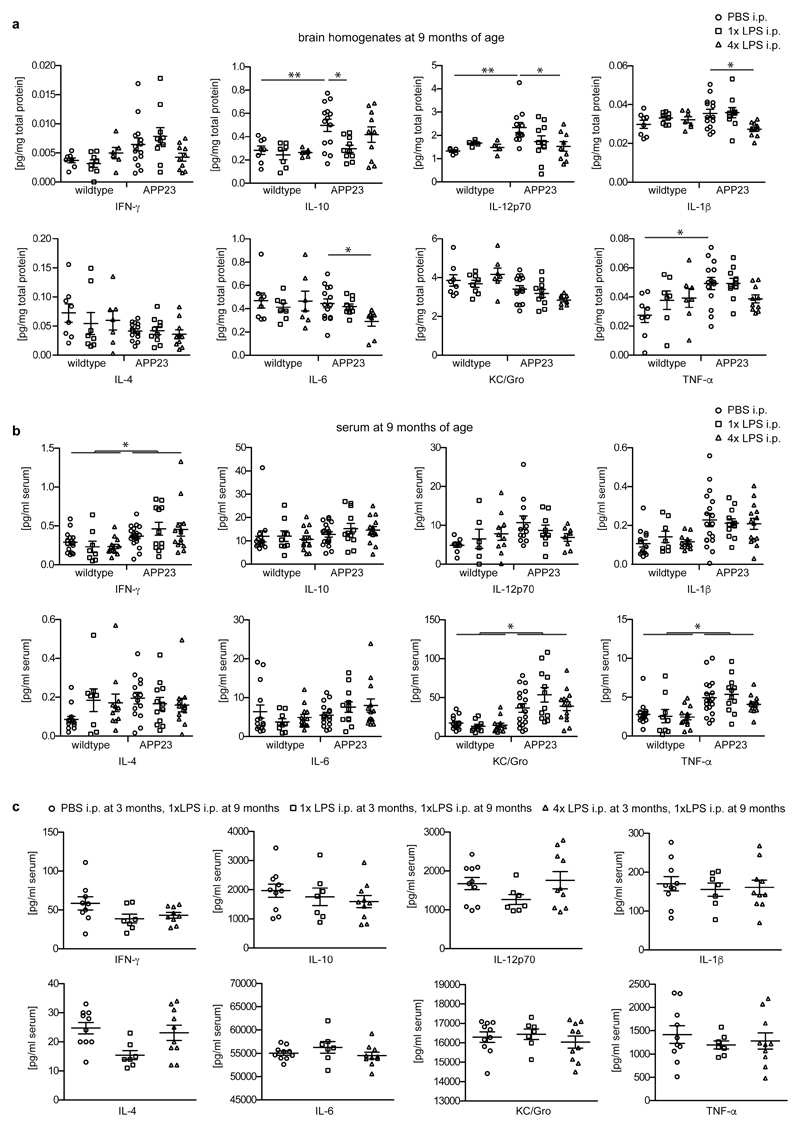
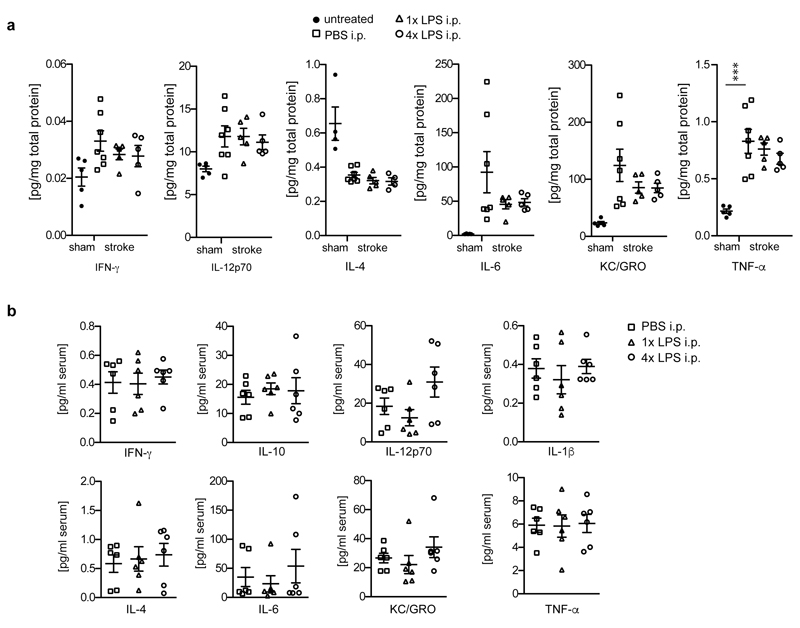
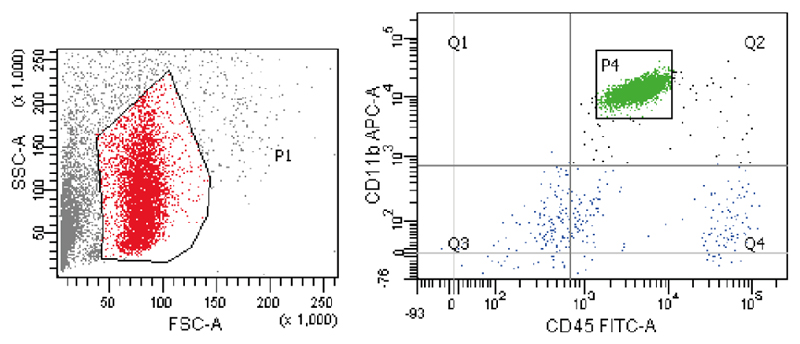
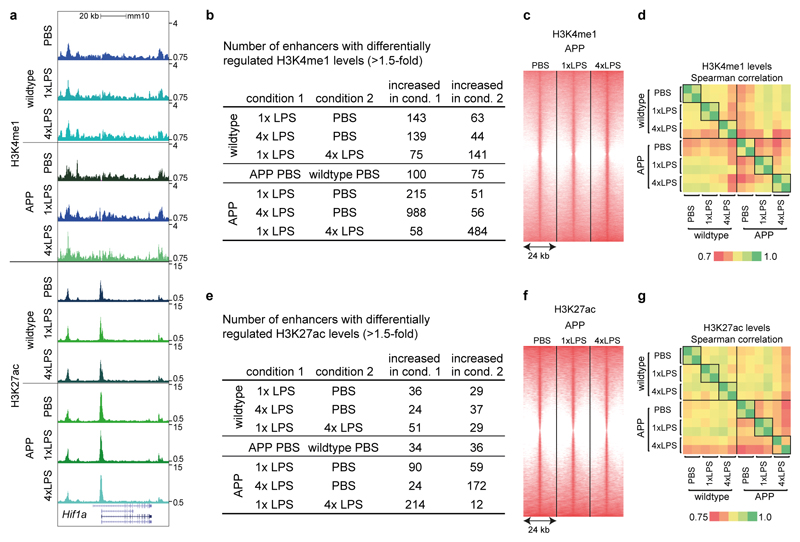
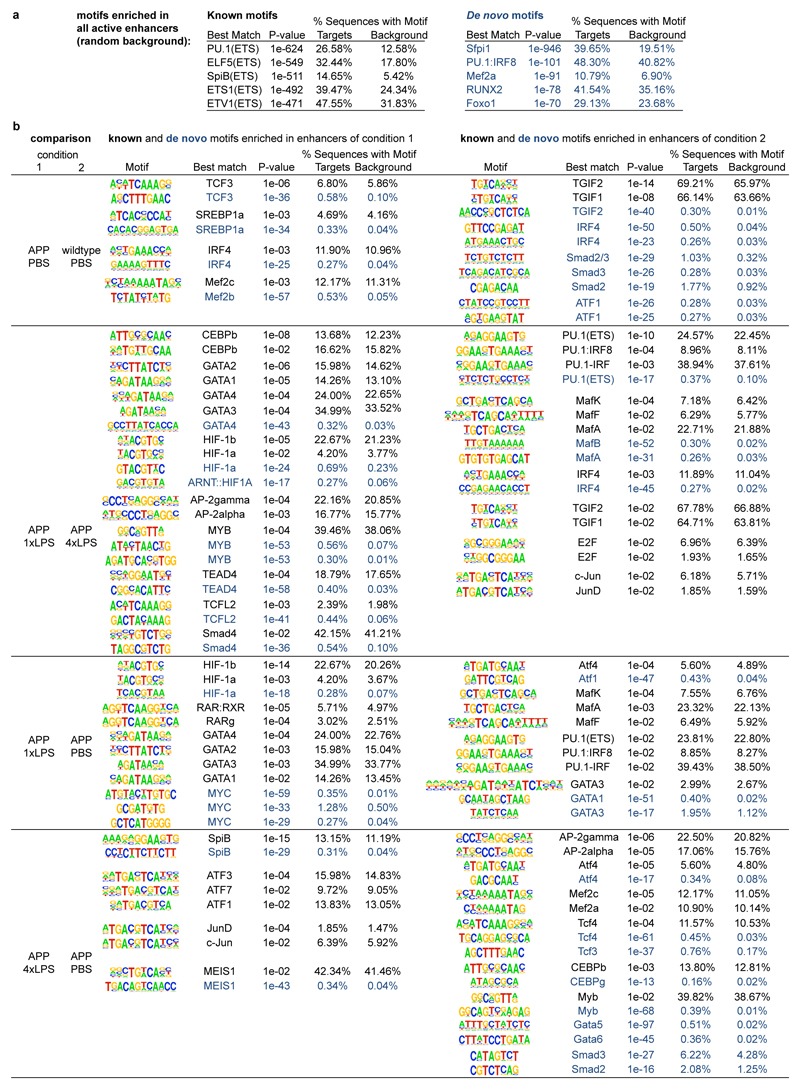
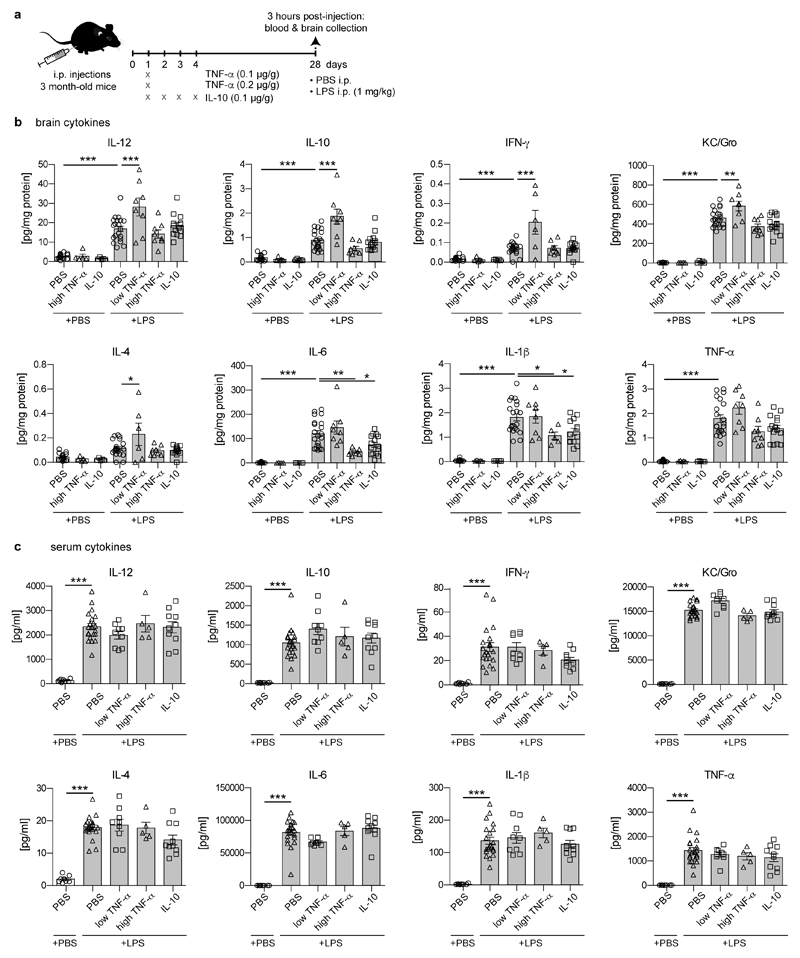
Comment in
-
Immune memory in the brain.Nature. 2018 Apr;556(7701):312-313. doi: 10.1038/d41586-018-03800-6. Nature. 2018. PMID: 29662132 No abstract available.
-
Microglial memory.Nat Rev Immunol. 2018 Jun;18(6):358-359. doi: 10.1038/s41577-018-0015-5. Nat Rev Immunol. 2018. PMID: 29713017 No abstract available.
-
Trained Microglia Trigger Memory Loss.Immunity. 2018 May 15;48(5):849-851. doi: 10.1016/j.immuni.2018.04.033. Immunity. 2018. PMID: 29768172
-
Training neuropathology.Nat Med. 2018 Jun;24(6):702. doi: 10.1038/s41591-018-0076-9. Nat Med. 2018. PMID: 29875460 No abstract available.
References
-
- Netea MG, Latz E, Mills KHG, O'Neill LAJ. Innate immune memory: a paradigm shift in understanding host defense. Nature Immunology. 2015;16:675–679. - PubMed
-
- Biswas SK, Lopez-Collazo E. Endotoxin tolerance: new mechanisms, molecules and clinical significance. Trends in Immunology. 2009;30:475–487. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
