

River networks as ecological corridors: A coherent ecohydrological perspective
- PMID: 29651194
- PMCID: PMC5890385
- DOI: 10.1016/j.advwatres.2017.10.005
River networks as ecological corridors: A coherent ecohydrological perspective
Abstract
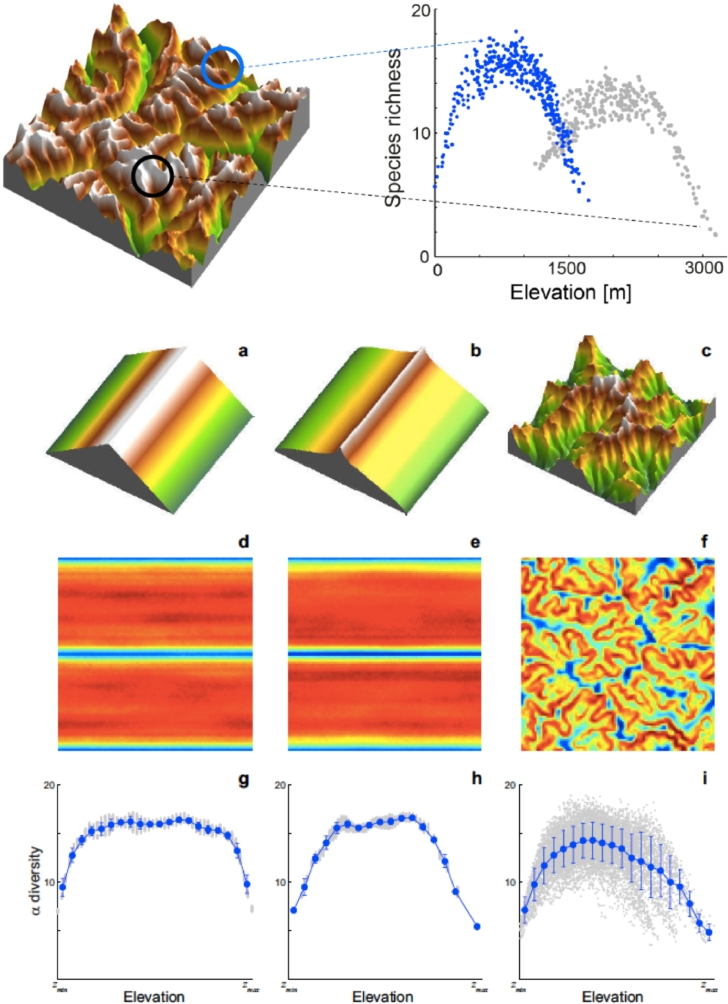
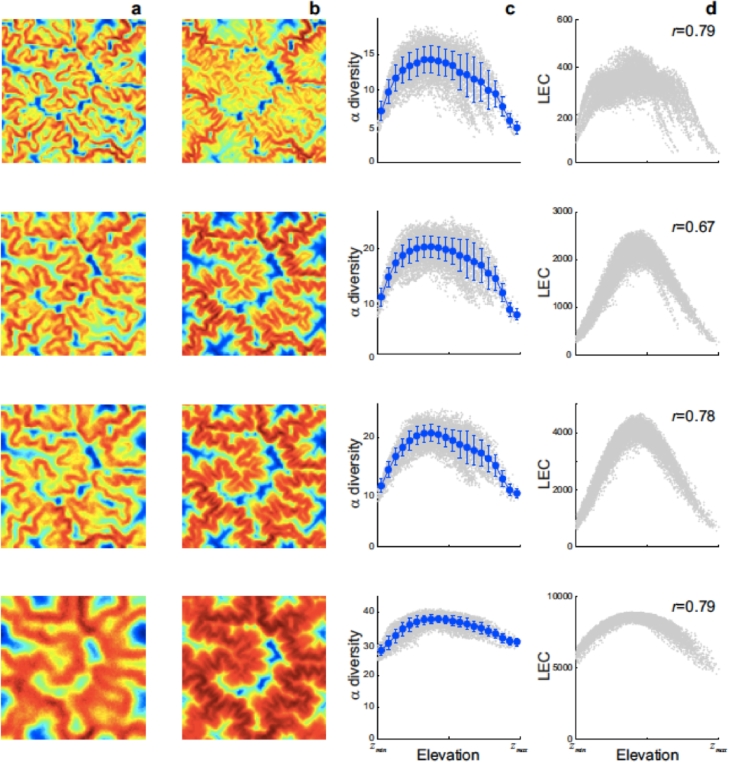
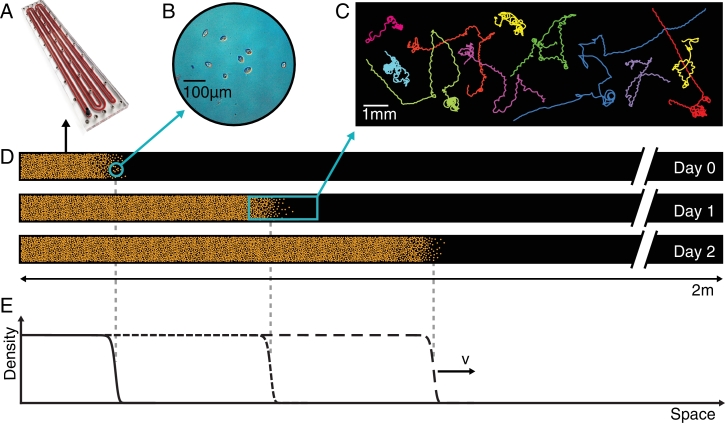
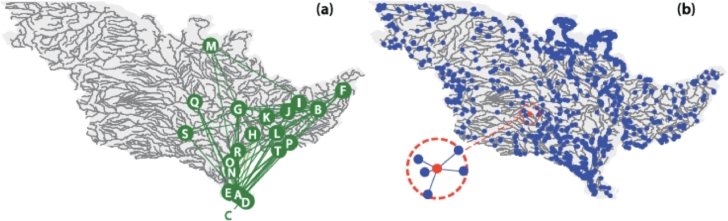
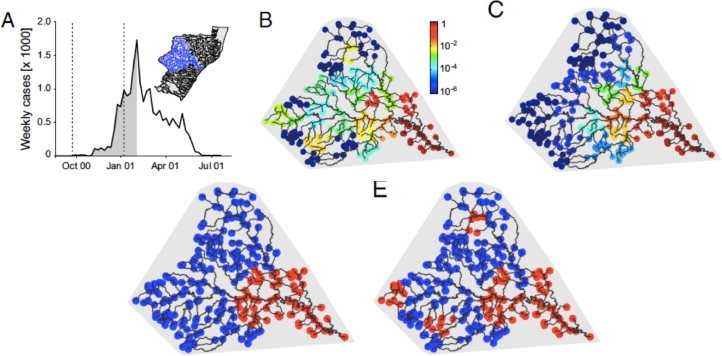
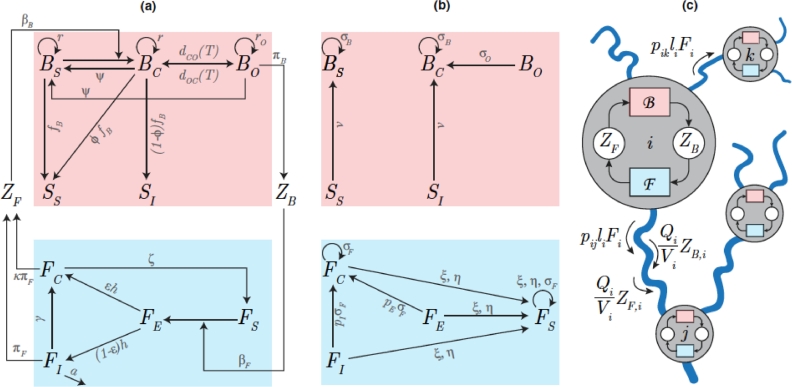
This paper draws together several lines of argument to suggest that an ecohydrological framework, i.e. laboratory, field and theoretical approaches focused on hydrologic controls on biota, has contributed substantially to our understanding of the function of river networks as ecological corridors. Such function proves relevant to: the spatial ecology of species; population dynamics and biological invasions; the spread of waterborne disease. As examples, we describe metacommunity predictions of fish diversity patterns in the Mississippi-Missouri basin, geomorphic controls imposed by the fluvial landscape on elevational gradients of species' richness, the zebra mussel invasion of the same Mississippi-Missouri river system, and the spread of proliferative kidney disease in salmonid fish. We conclude that spatial descriptions of ecological processes in the fluvial landscape, constrained by their specific hydrologic and ecological dynamics and by the ecosystem matrix for interactions, i.e. the directional dispersal embedded in fluvial and host/pathogen mobility networks, have already produced a remarkably broad range of significant results. Notable scientific and practical perspectives are thus open, in the authors' view, to future developments in ecohydrologic research.
Keywords: Directional dispersal; Metacommunity models; Metapopulation models; Spatially explicit ecology; Substrate topology.
Figures
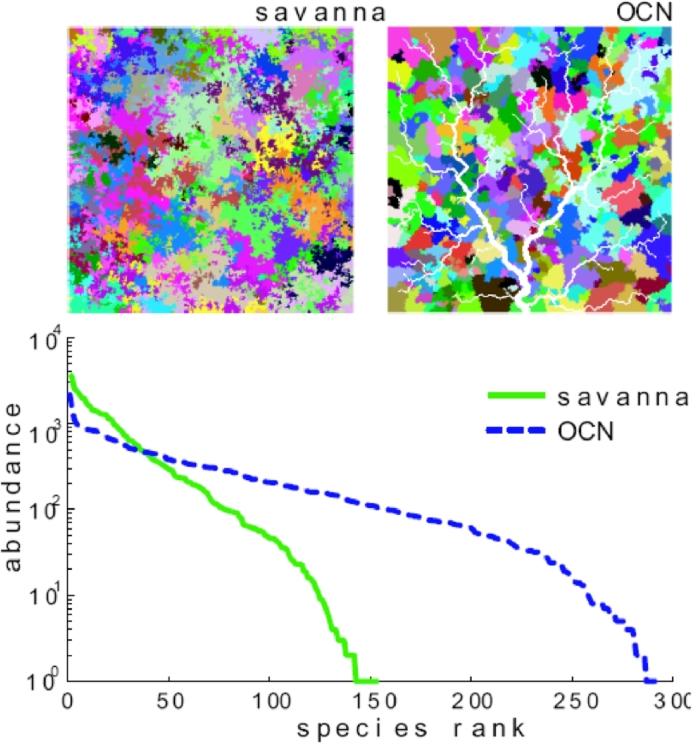
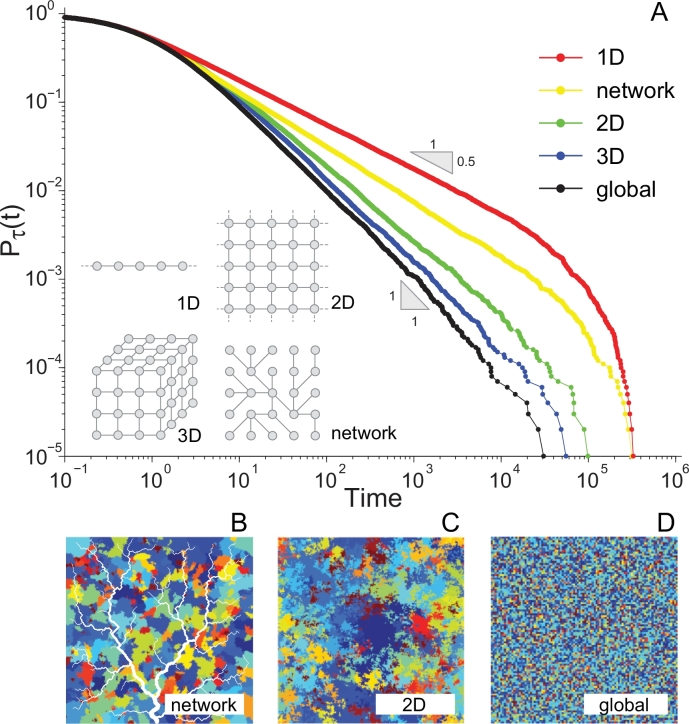


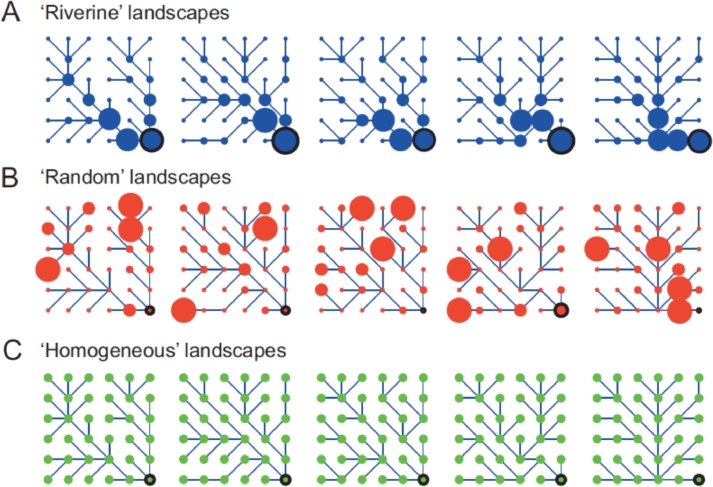
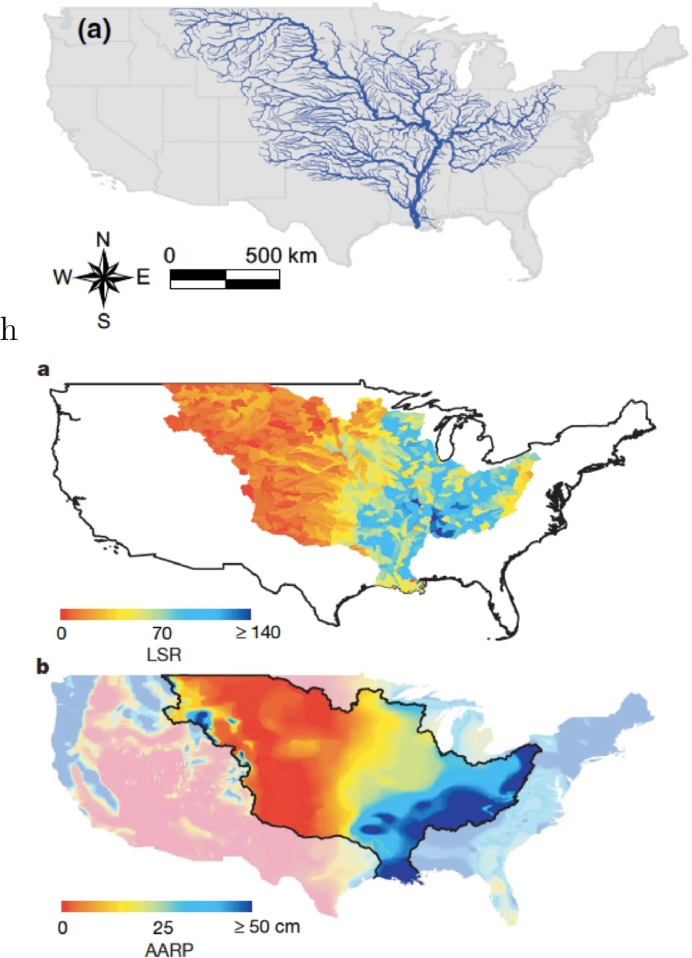










References
-
- de Aguiar M., Baranger M., Baptestini E.M., Kaufman L., Bar-Yam Y. Global patterns of speciation and diversity. Nature. 2009;460(7253):384–388. - PubMed
-
- Akaike H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1974;19:716–723.
-
- Alexander R., Jones P., Boyer E., Smith R. Effect of stream channel size on the delivery of nitrogen to the Gulf of Mexico. Nature. 2000;403:758–761. - PubMed
-
- Allen Y.C., Ramcharan C. Dreissena distribution in commercial waterways of the US: using failed invasions to identify limiting factors. Can. J. Fish. Aquat. Sci. 2001;58:898–907.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials