

FAR2 is associated with kidney disease in mice and humans
- PMID: 29652635
- PMCID: PMC6139637
- DOI: 10.1152/physiolgenomics.00118.2017
FAR2 is associated with kidney disease in mice and humans
Abstract
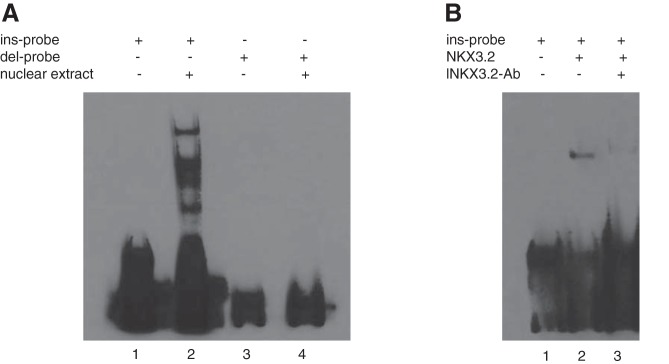
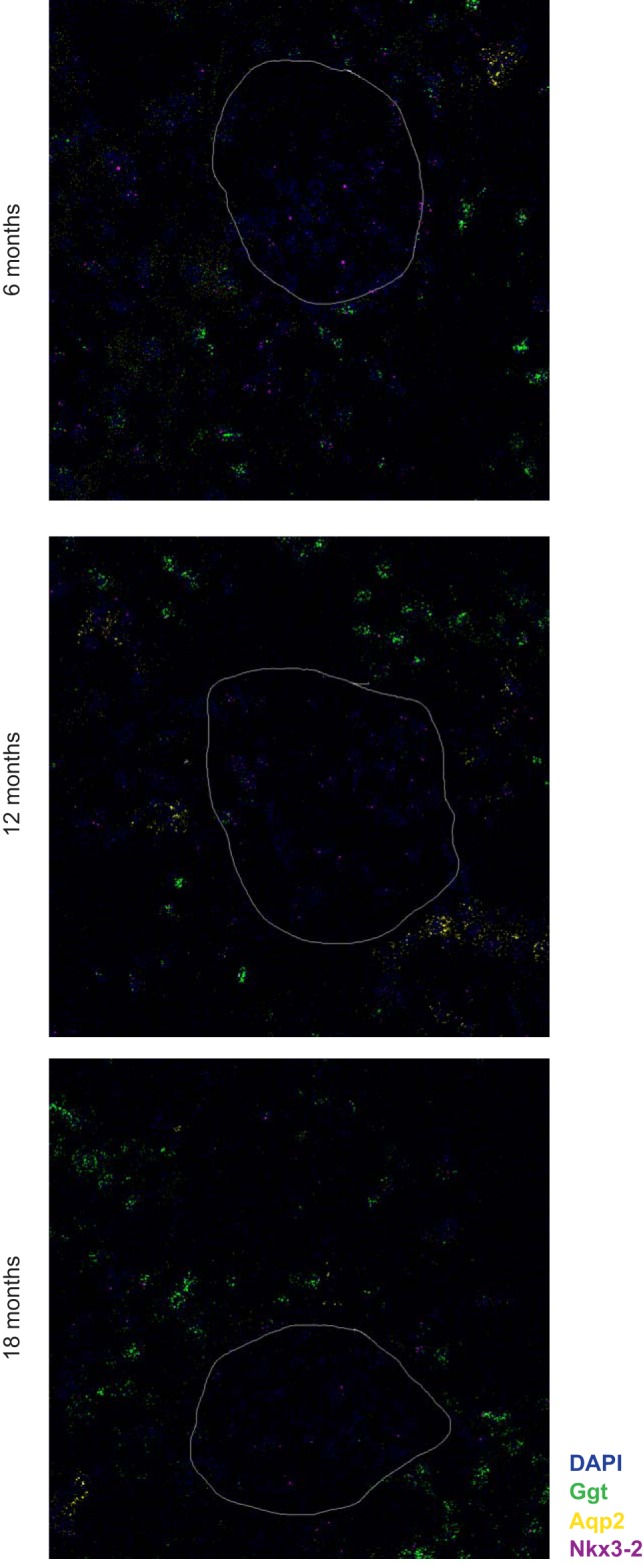
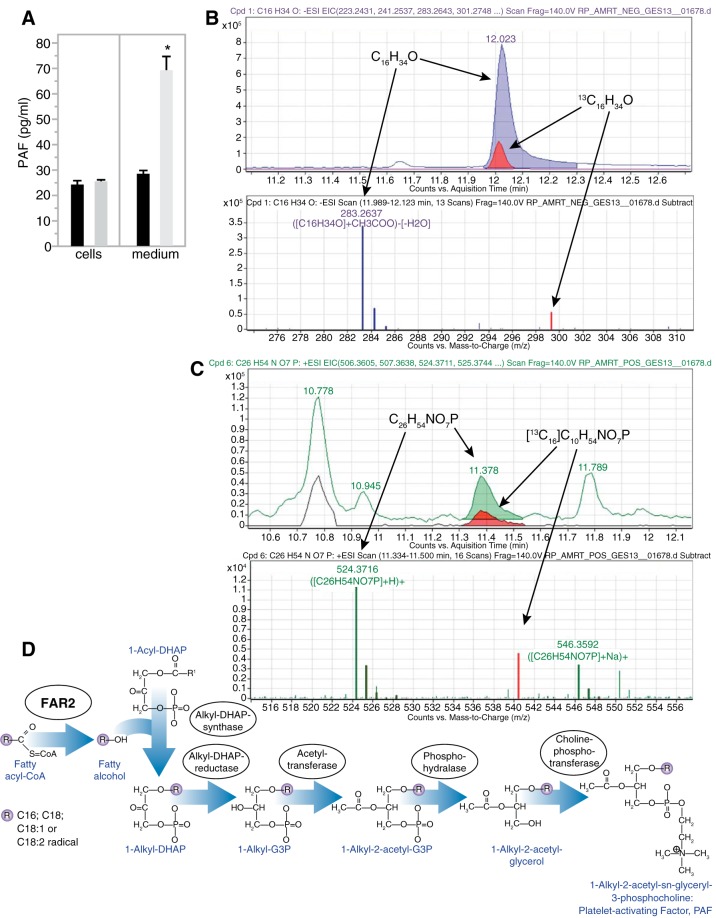
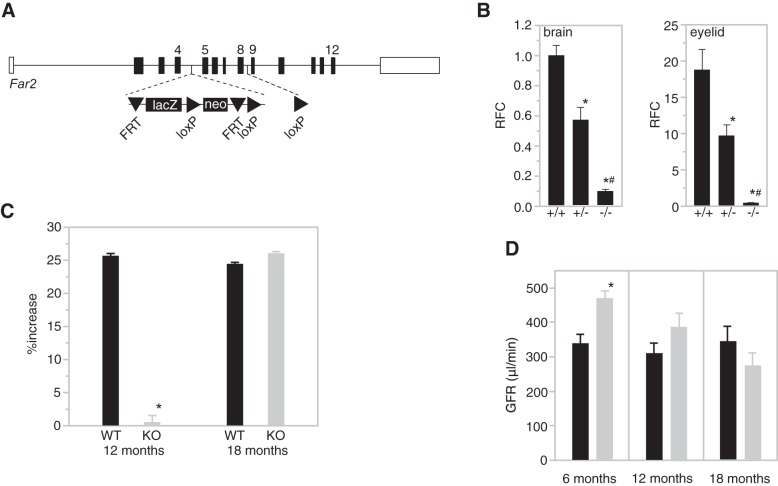
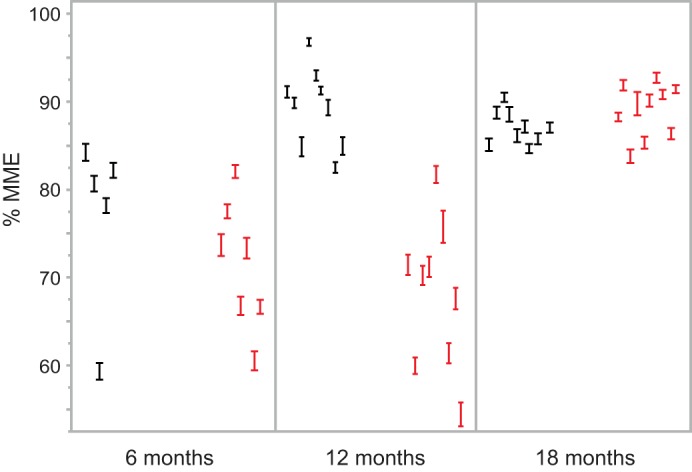
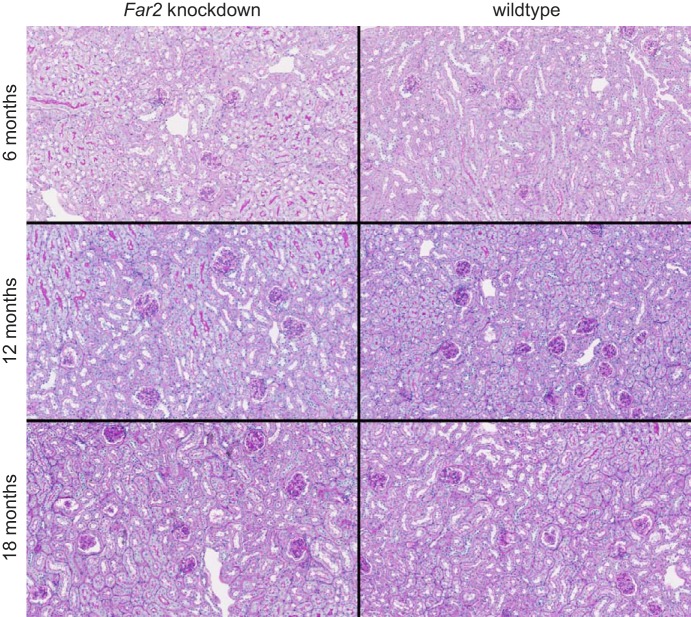
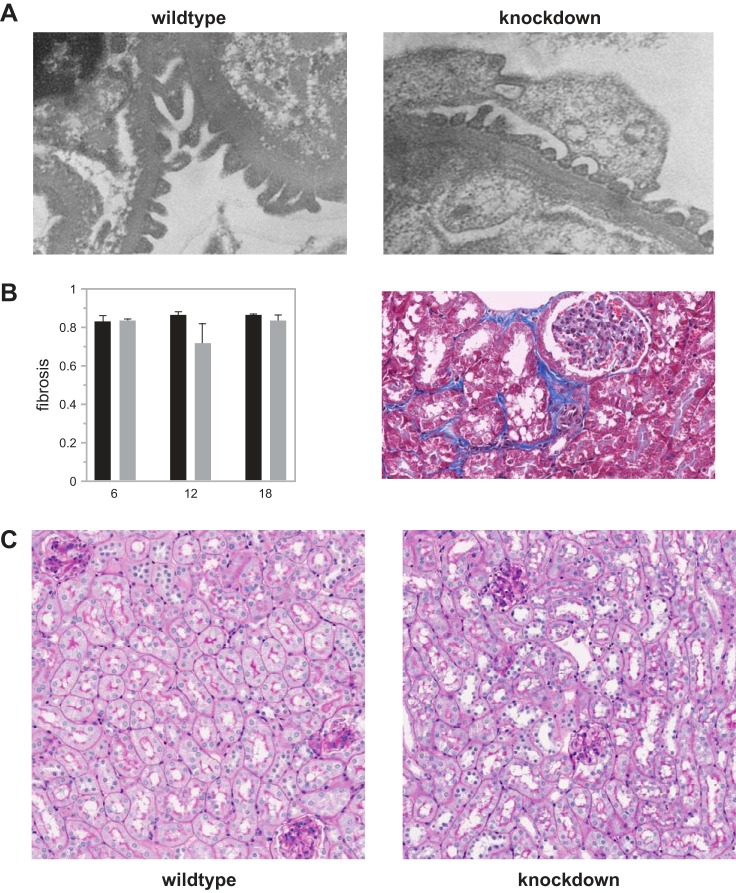
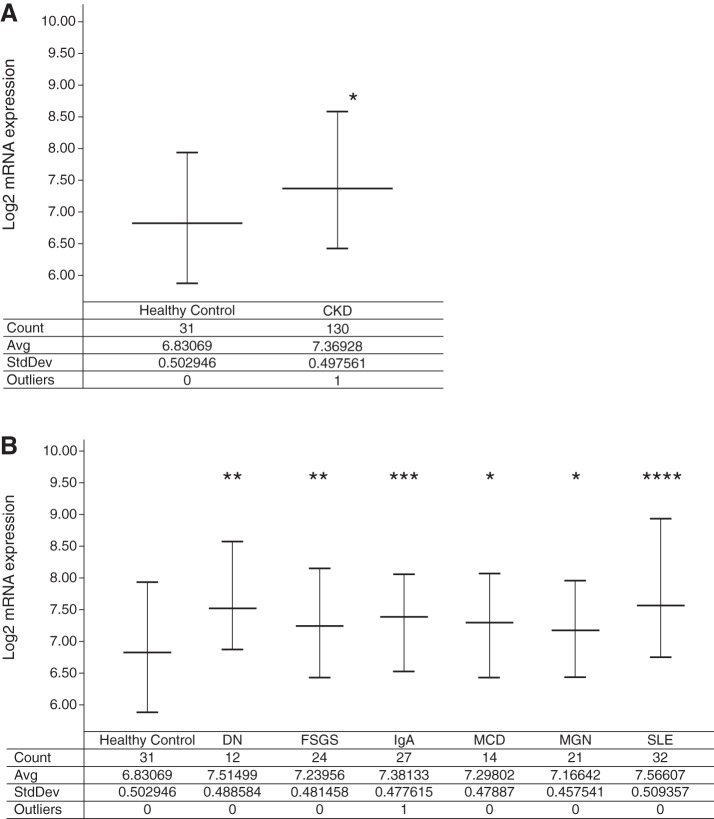
Mesangial matrix expansion is an important process in the initiation of chronic kidney disease, yet the genetic factors driving its development are unknown. Our previous studies have implicated Far2 as a candidate gene associated with differences in mesangial matrix expansion between mouse inbred strains. Consistent with the hypothesis that increased expression of Far2 leads to mesangial matrix expansion through increased production of platelet-activating factor precursors, we show that FAR2 is capable of mediating de novo platelet-activating factor synthesis in vitro and driven by the transcription factor NKX3.2. We demonstrate that knockdown of Far2 in mice delays the progression of mesangial matrix expansion with at least six months (equivalent to ~15 yr in human). Furthermore, we show that increased FAR2 expression in human patients is associated with diabetic nephropathy, lupus nephritis, and IgA nephropathy. Taken together, these results highlight FAR2's role in the development of mesangial matrix expansion and chronic kidney disease.
Keywords: aging; kidney; mesangial matrix; mouse.
Figures
References
-
- Barrett T, Wilhite SE, Ledoux P, Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH, Sherman PM, Holko M, Yefanov A, Lee H, Zhang N, Robertson CL, Serova N, Davis S, Soboleva A. NCBI GEO: archive for functional genomics data sets–update. Nucleic Acids Res 41, D1: D991–D995, 2013. doi:10.1093/nar/gks1193. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous