

Water-stress induced downsizing of light-harvesting antenna complex protects developing rice seedlings from photo-oxidative damage
- PMID: 29654242
- PMCID: PMC5899091
- DOI: 10.1038/s41598-017-14419-4
Water-stress induced downsizing of light-harvesting antenna complex protects developing rice seedlings from photo-oxidative damage
Abstract
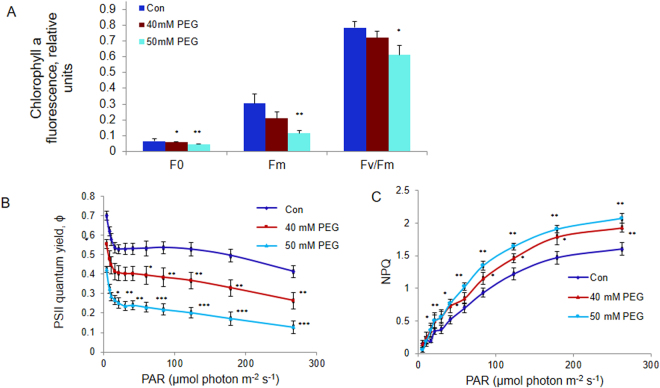
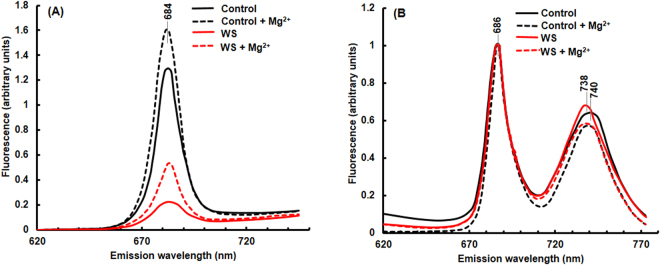
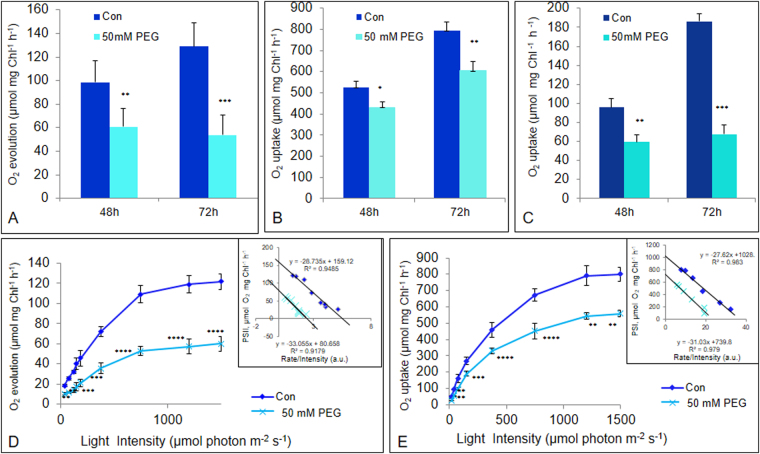
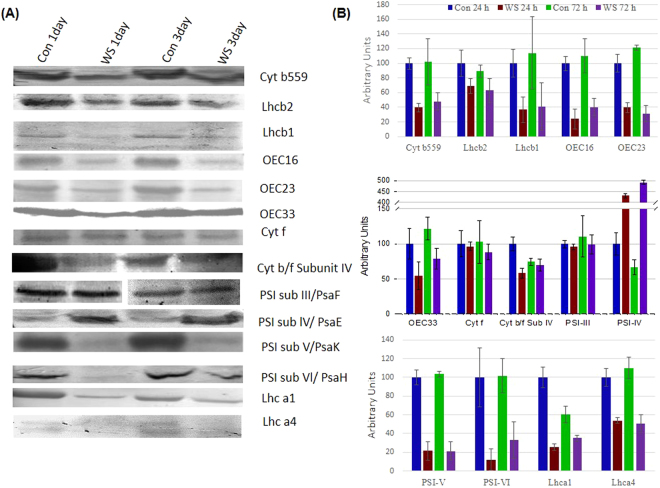
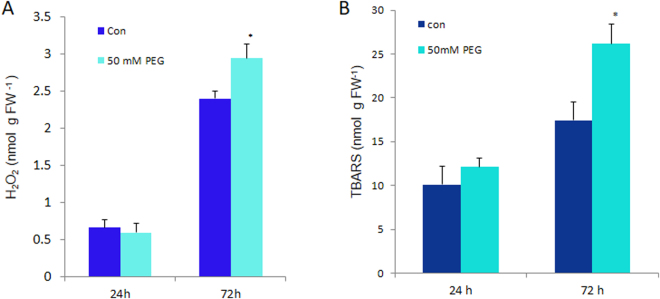
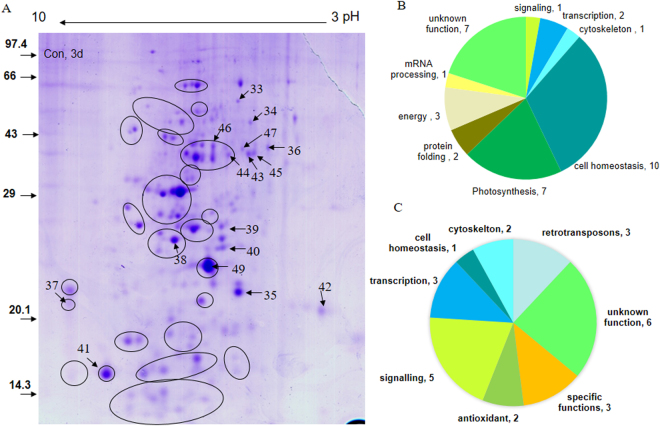
The impact of water-stress on chloroplast development was studied by applying polyethylene glycol 6000 to the roots of 5-day-old etiolated rice (Oryza sativa) seedlings that were subsequently illuminated up to 72 h. Chloroplast development in drought environment led to down-regulation of light-harvesting Chl-proteins. Photosynthetic proteins of Photosystem II (PSII) and oxygen evolving complex i.e., Cytb559, OEC16, OEC23 and OEC33 as well as those of PSI such as PSI-III, PSI-V, and PSI-VI, decreased in abundance. Consequently, due to reduced light absorption by antennae, the electron transport rates of PSII and PSI decreased by 55% and 25% respectively. Further, seedling development in stress condition led to a decline in the ratio of variable (Fv) to maximum (Fm) Chl a fluorescence, as well in the quantum yield of PSII photochemistry. Addition of Mg2+ to the thylakoid membranes suggested that Mg2+-induced grana stacking was not affected by water deficit. Proteomic analysis revealed the down-regulation of proteins involved in electron transport and in carbon reduction reactions, and up-regulation of antioxidative enzymes. Our results demonstrate that developing seedlings under water deficit could downsize their light-harvesting capacity and components of photosynthetic apparatus to prevent photo-oxidative stress, excess ROS generation and membrane lipid peroxidation.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Pareek, A., Sopory, S. K. & Bohnert, H. J. Govindjee (eds) Abiotic Stress Adaptation in Plants, Physiological, Molecular and Genomic Foundation (Springer, Dordrecht, 2010).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
