

Photoreceptor-induced RPE phagolysosomal maturation defects in Stargardt-like Maculopathy (STGD3)
- PMID: 29654292
- PMCID: PMC5899129
- DOI: 10.1038/s41598-018-24357-4
Photoreceptor-induced RPE phagolysosomal maturation defects in Stargardt-like Maculopathy (STGD3)
Abstract
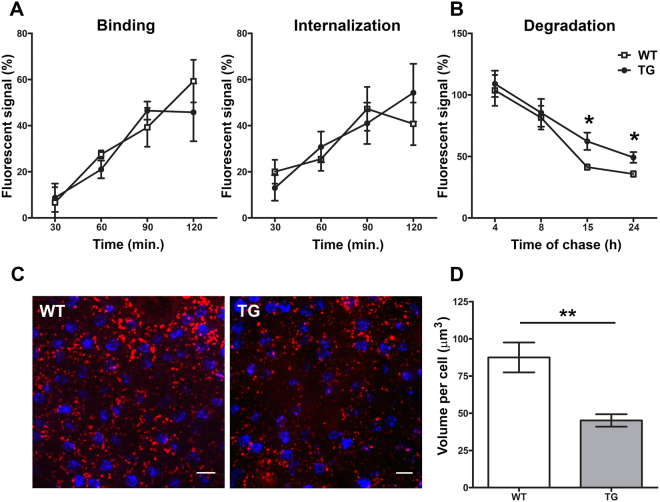
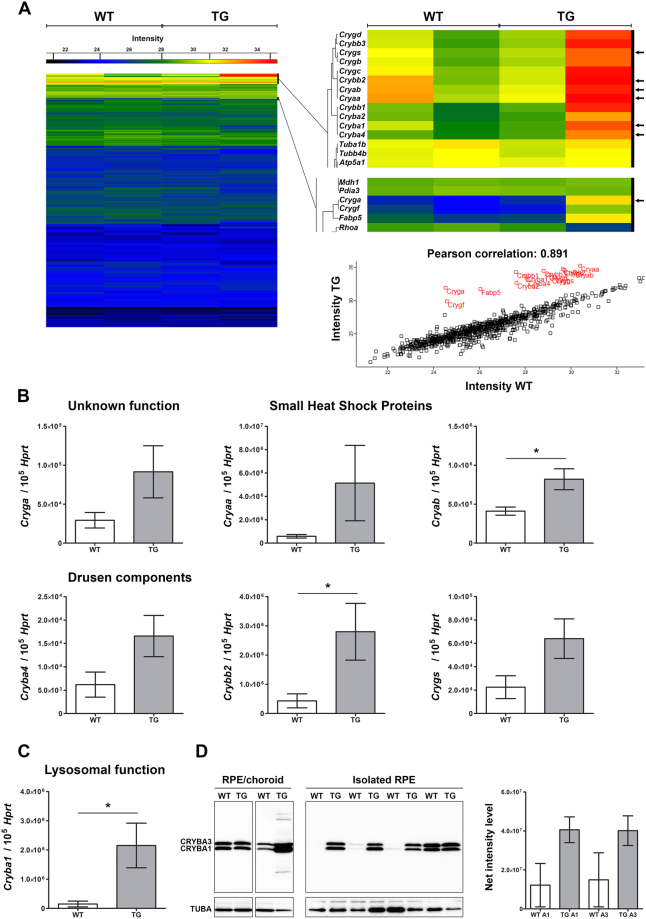
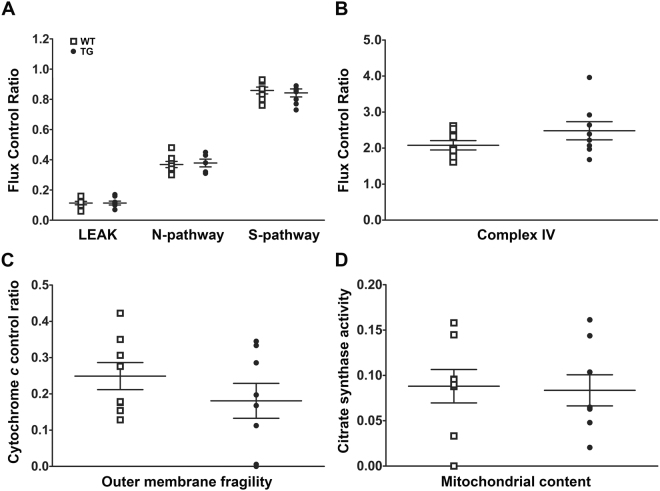
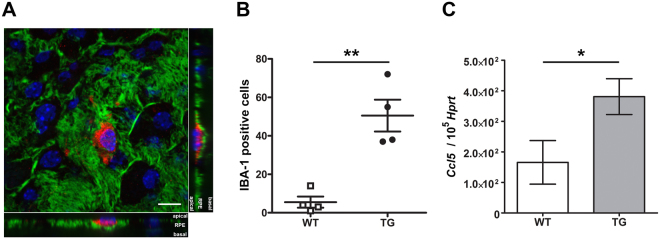
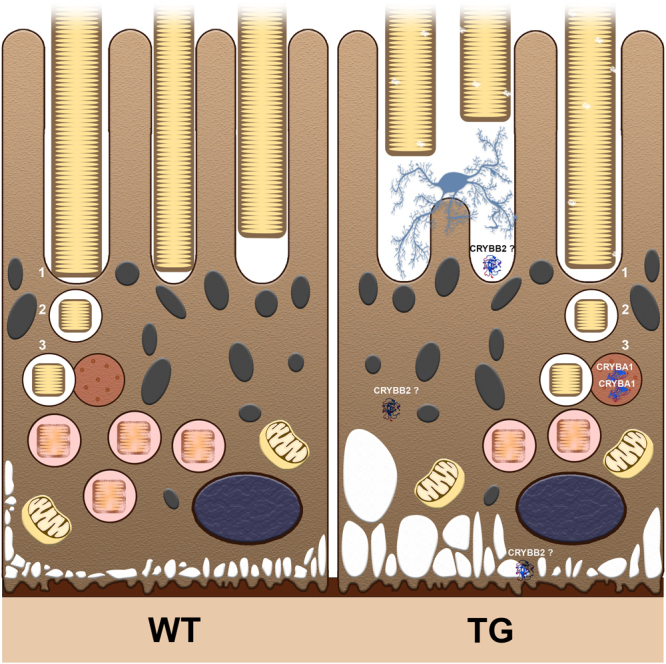
For many neurodegenerative disorders, expression of a pathological protein by one cell type impedes function of other cell types, which in turn contributes to the death of the first cell type. In transgenic mice modelling Stargardt-like (STGD3) maculopathy, human mutant ELOVL4 expression by photoreceptors is associated with defects in the underlying retinal pigment epithelium (RPE). To examine how photoreceptors exert cytotoxic effects on RPE cells, transgenic ELOVL4 (TG1-2 line; TG) and wild-type (WT) littermates were studied one month prior (preclinical stage) to onset of photoreceptor loss (two months). TG photoreceptor outer segments presented to human RPE cells are recognized and internalized into phagosomes, but their digestion is delayed. Live RPE cell imaging pinpoints decreased numbers of acidified phagolysomes. In vivo, master regulator of lysosomal genes, transcription factor EB (TFEB), and key lysosomal enzyme Cathepsin D are both unaffected. Oxidative stress, as ruled out with high-resolution respirometry, does not play a role at such an early stage. Upregulation of CRYBA1/A3 and phagocytic cells (microglia/macrophages) interposed between RPE and photoreceptors support adaptive responses to processing delays. Impaired phagolysosomal maturation is observed in RPE of mice expressing human mutant ELOVL4 in their photoreceptors prior to photoreceptor death and associated vision loss.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
