

Structural Mechanism of Functional Modulation by Gene Splicing in NMDA Receptors
- PMID: 29656875
- PMCID: PMC5963293
- DOI: 10.1016/j.neuron.2018.03.034
Structural Mechanism of Functional Modulation by Gene Splicing in NMDA Receptors
Abstract
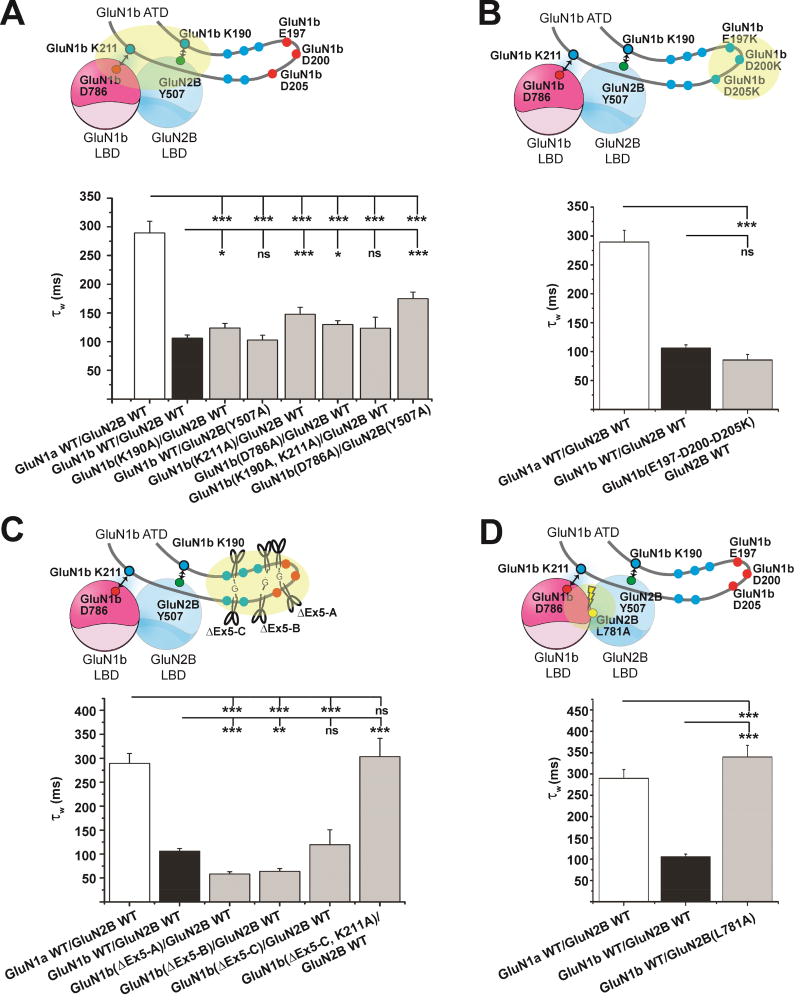
Alternative gene splicing gives rise to N-methyl-D-aspartate (NMDA) receptor ion channels with defined functional properties and unique contributions to calcium signaling in a given chemical environment in the mammalian brain. Splice variants possessing the exon-5-encoded motif at the amino-terminal domain (ATD) of the GluN1 subunit are known to display robustly altered deactivation rates and pH sensitivity, but the underlying mechanism for this functional modification is largely unknown. Here, we show through cryoelectron microscopy (cryo-EM) that the presence of the exon 5 motif in GluN1 alters the local architecture of heterotetrameric GluN1-GluN2 NMDA receptors and creates contacts with the ligand-binding domains (LBDs) of the GluN1 and GluN2 subunits, which are absent in NMDA receptors lacking the exon 5 motif. The unique interactions established by the exon 5 motif are essential to the stability of the ATD/LBD and LBD/LBD interfaces that are critically involved in controlling proton sensitivity and deactivation.
Keywords: NMDA receptor; alternative splicing; cryo-EM; electrophysiology.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Authors declare no conflict of interests.
Figures





References
-
- Adams DR, Yuan H, Holyoak T, Arajs KH, Hakimi P, Markello TC, Wolfe LA, Vilboux T, Burton BK, Fajardo KF, et al. Three rare diseases in one Sib pair: RAI1, PCK1, GRIN2B mutations associated with Smith-Magenis Syndrome, cytosolic PEPCK deficiency and NMDA receptor glutamate insensitivity. Molecular genetics and metabolism. 2014;113:161–170. - PMC - PubMed
-
- Afonine PV, Headd JJ, Terwilliger TC, Adams PD. Computational Crystallography Newsletter. 2013;4:43–44.
-
- Bassler EL, Ngo-Anh TJ, Geisler HS, Ruppersberg JP, Grunder S. Molecular and functional characterization of acid-sensing ion channel (ASIC) 1b. J Biol Chem. 2001;276:33782–33787. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
