

Warming Induces Significant Reprogramming of Beige, but Not Brown, Adipocyte Cellular Identity
- PMID: 29657031
- PMCID: PMC5932137
- DOI: 10.1016/j.cmet.2018.03.005
Warming Induces Significant Reprogramming of Beige, but Not Brown, Adipocyte Cellular Identity
Abstract
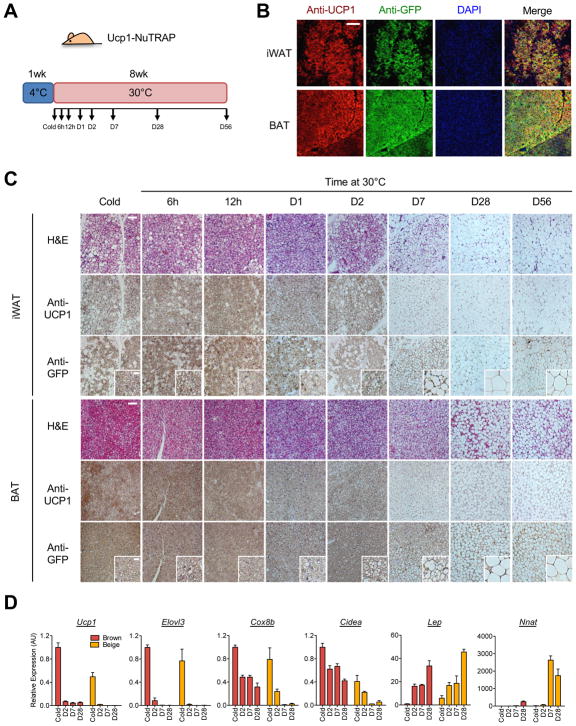
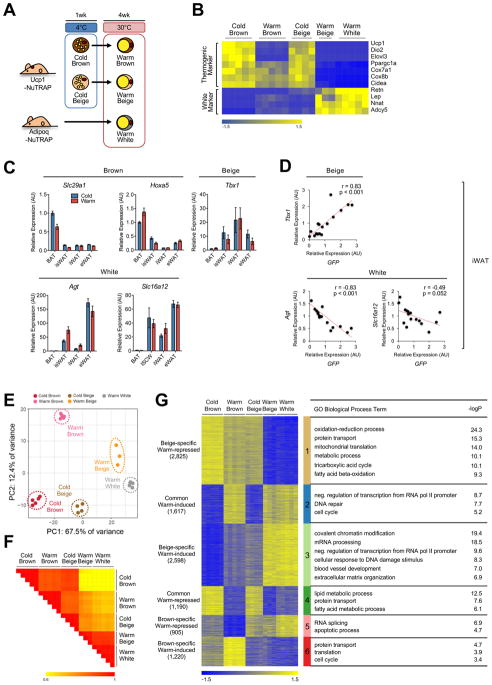
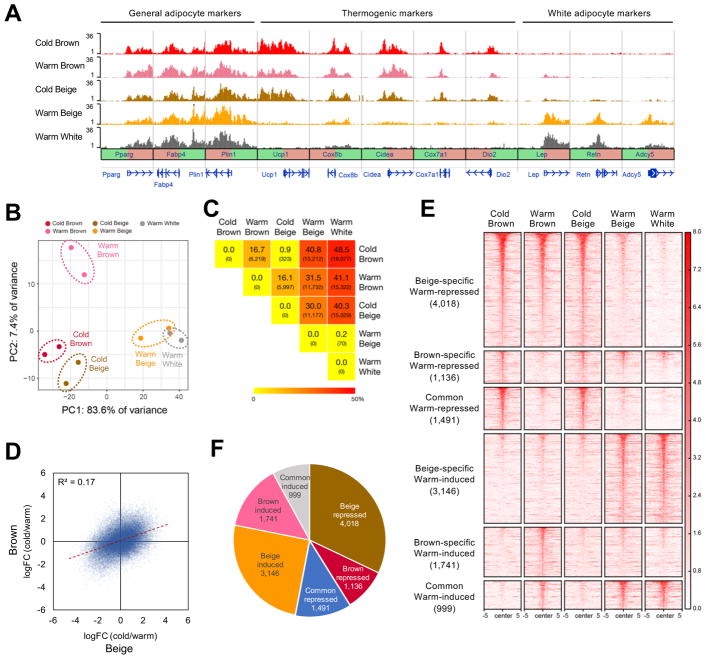
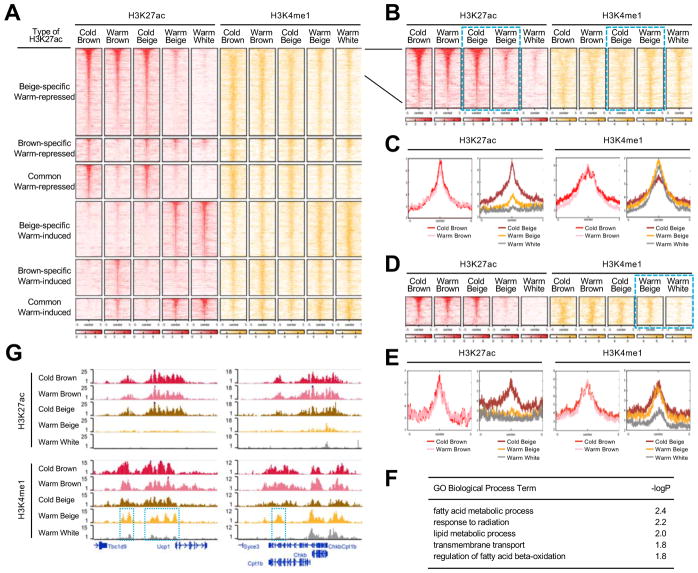
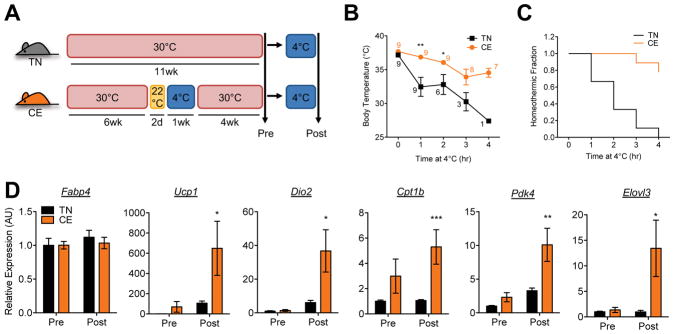
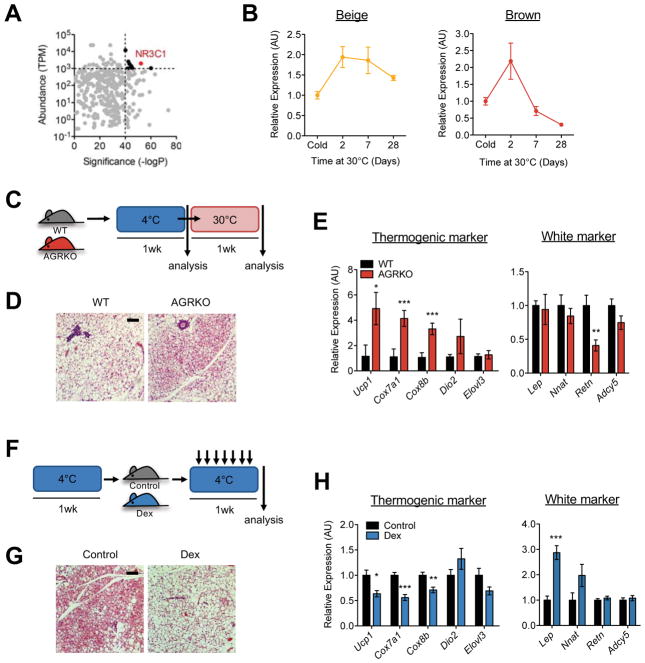
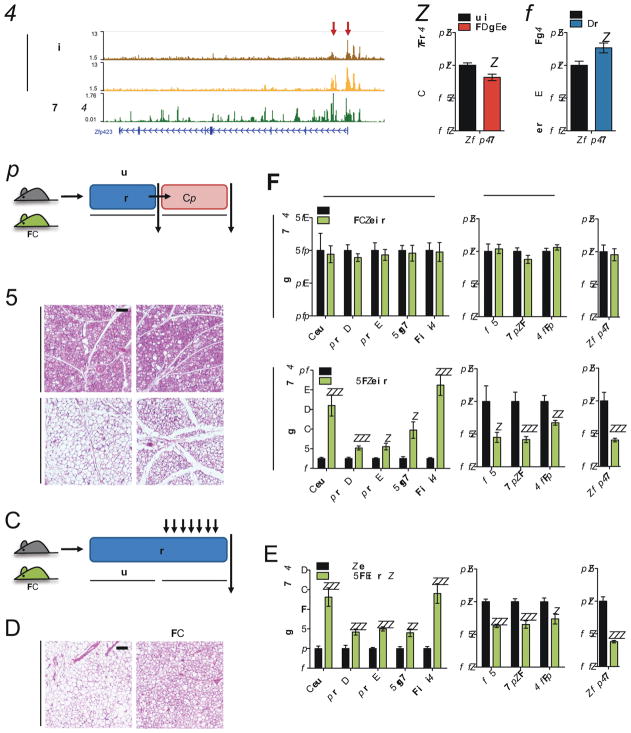
Beige and brown adipocytes generate heat in response to reductions in ambient temperature. When warmed, both beige and brown adipocytes exhibit morphological "whitening," but it is unknown whether or to what extent this represents a true shift in cellular identity. Using cell-type-specific profiling in vivo, we uncover a unique paradigm of temperature-dependent epigenomic plasticity of beige, but not brown, adipocytes, with conversion from a brown to a white chromatin state. Despite this profound shift in cellular identity, warm whitened beige adipocytes retain an epigenomic memory of prior cold exposure defined by an array of poised enhancers that prime thermogenic genes for rapid response during a second bout of cold exposure. We further show that a transcriptional cascade involving glucocorticoid receptor and Zfp423 can drive warm-induced whitening of beige adipocytes. These studies identify the epigenomic and transcriptional bases of an extraordinary example of cellular plasticity in response to environmental signals.
Keywords: Zfp423; adipocyte; beige; brown; epigenome; epigenomic memory; glucocorticoid receptor; reprogramming; transcriptome; white.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
These authors have no competing interests to declare.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
