

Functional Maps of Mechanosensory Features in the Drosophila Brain
- PMID: 29657118
- PMCID: PMC5952606
- DOI: 10.1016/j.cub.2018.02.074
Functional Maps of Mechanosensory Features in the Drosophila Brain
Abstract
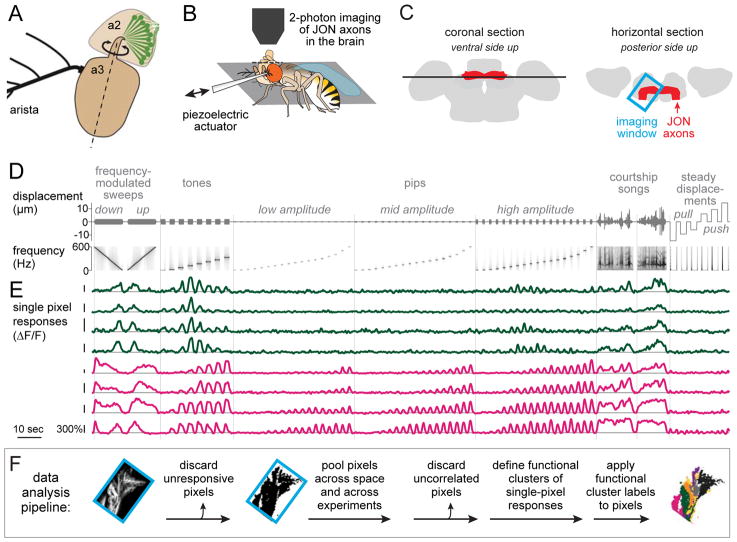
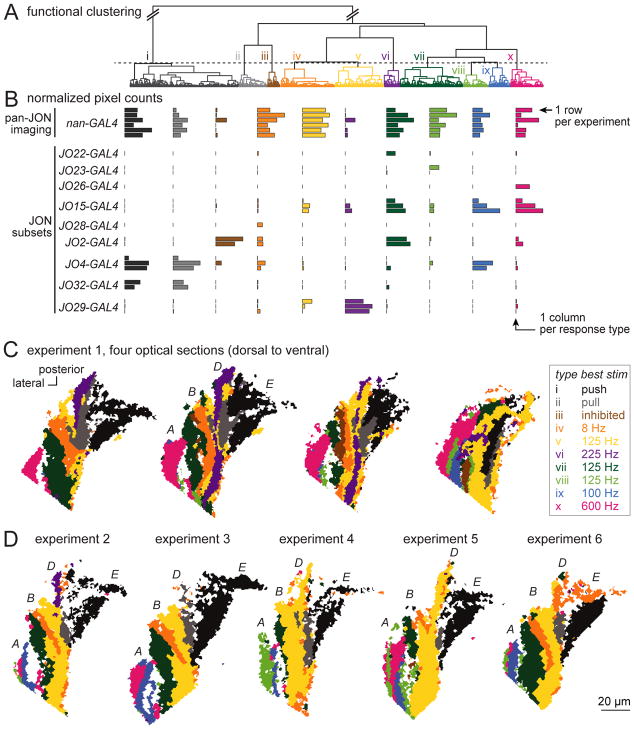
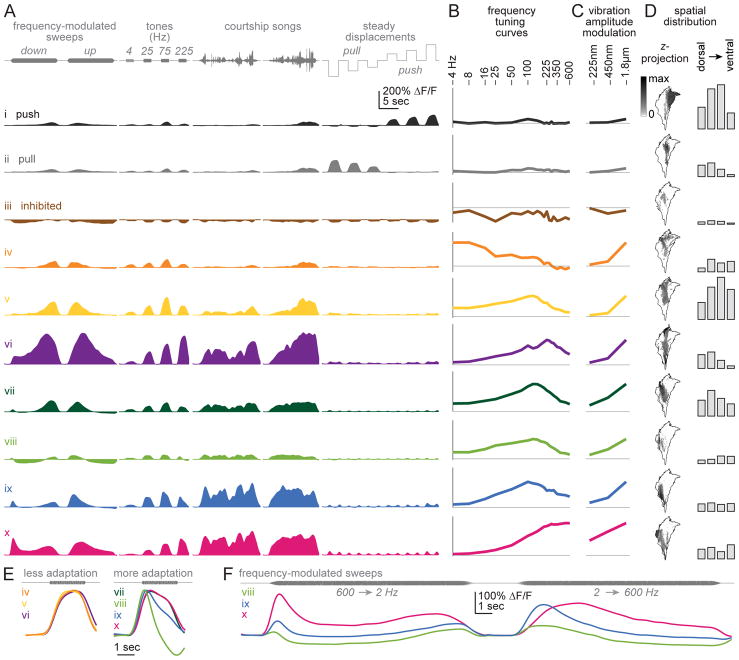
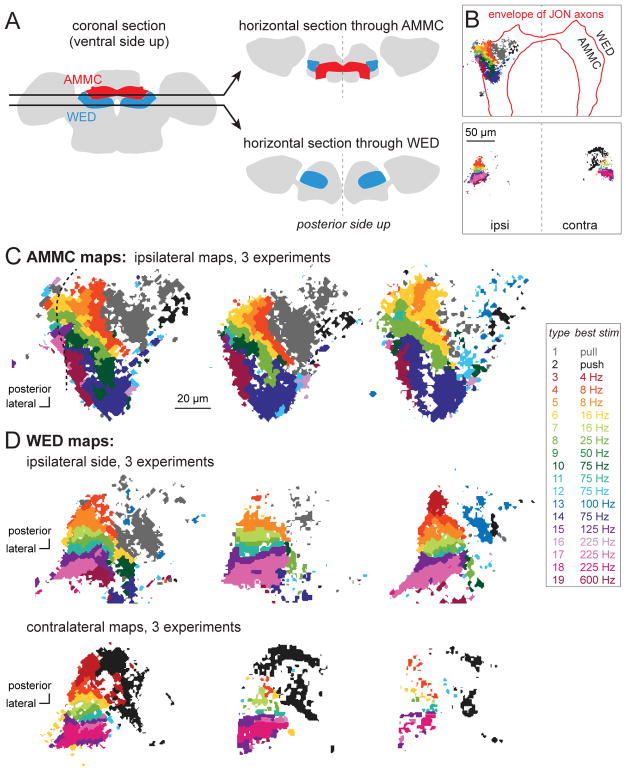
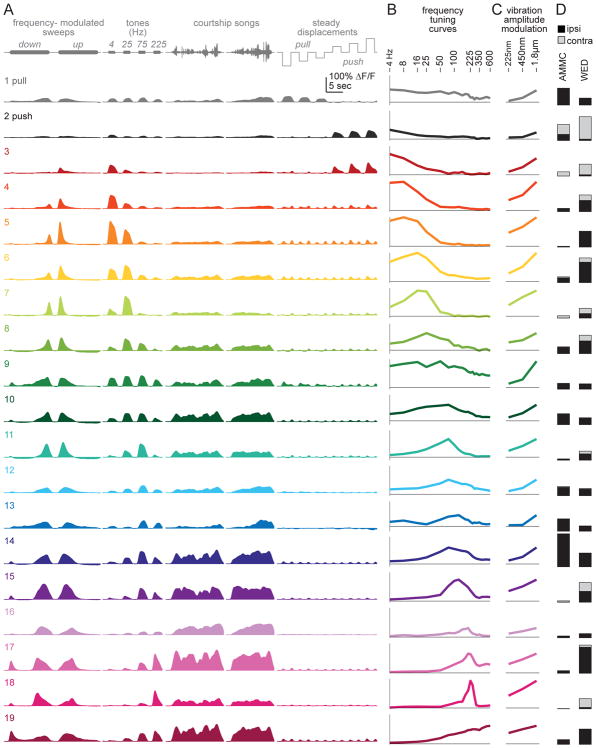
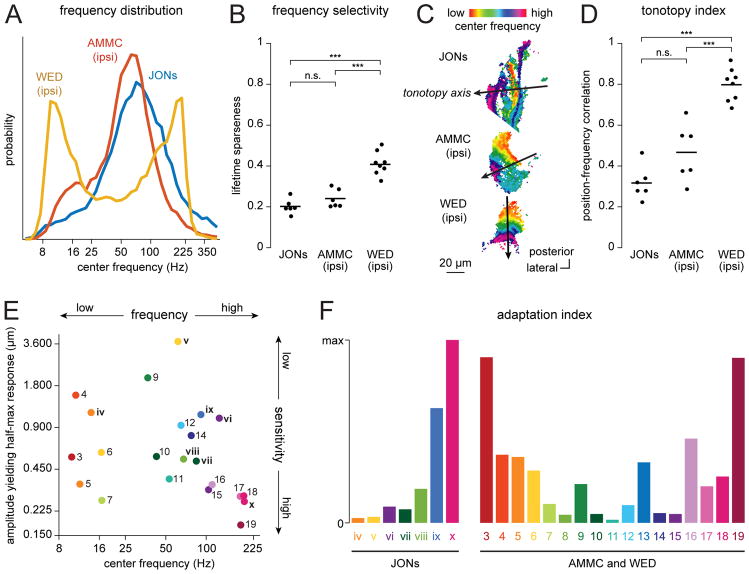
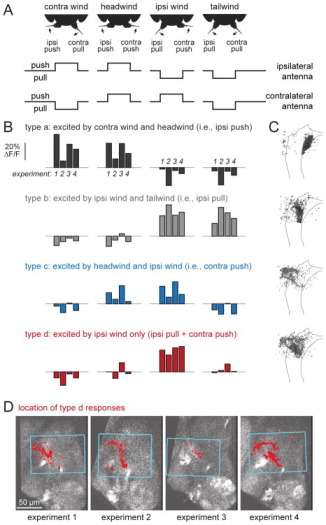
Johnston's organ is the largest mechanosensory organ in Drosophila. It contributes to hearing, touch, vestibular sensing, proprioception, and wind sensing. In this study, we used in vivo 2-photon calcium imaging and unsupervised image segmentation to map the tuning properties of Johnston's organ neurons (JONs) at the site where their axons enter the brain. We then applied the same methodology to study two key brain regions that process signals from JONs: the antennal mechanosensory and motor center (AMMC) and the wedge, which is downstream of the AMMC. First, we identified a diversity of JON response types that tile frequency space and form a rough tonotopic map. Some JON response types are direction selective; others are specialized to encode amplitude modulations over a specific range (dynamic range fractionation). Next, we discovered that both the AMMC and the wedge contain a tonotopic map, with a significant increase in tonotopy-and a narrowing of frequency tuning-at the level of the wedge. Whereas the AMMC tonotopic map is unilateral, the wedge tonotopic map is bilateral. Finally, we identified a subregion of the AMMC/wedge that responds preferentially to the coherent rotation of the two mechanical organs in the same angular direction, indicative of oriented steady air flow (directional wind). Together, these maps reveal the broad organization of the primary and secondary mechanosensory regions of the brain. They provide a framework for future efforts to identify the specific cell types and mechanisms that underlie the hierarchical re-mapping of mechanosensory information in this system.
Keywords: WED; auditory; chordotonal; hearing; phase; selectivity; sound; vibration.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Comprehensive classification of the auditory sensory projections in the brain of the fruit fly Drosophila melanogaster.J Comp Neurol. 2006 Nov 20;499(3):317-56. doi: 10.1002/cne.21075. J Comp Neurol. 2006. PMID: 16998934
-
Distinct sensory representations of wind and near-field sound in the Drosophila brain.Nature. 2009 Mar 12;458(7235):201-5. doi: 10.1038/nature07843. Nature. 2009. PMID: 19279637 Free PMC article.
-
GABAergic Local Interneurons Shape Female Fruit Fly Response to Mating Songs.J Neurosci. 2018 May 2;38(18):4329-4347. doi: 10.1523/JNEUROSCI.3644-17.2018. Epub 2018 Apr 24. J Neurosci. 2018. PMID: 29691331 Free PMC article.
-
Neuronal encoding of sound, gravity, and wind in the fruit fly.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2013 Apr;199(4):253-62. doi: 10.1007/s00359-013-0806-x. Epub 2013 Mar 13. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2013. PMID: 23494584 Review.
-
Hearing molecules, mechanism and transportation: modeled in Drosophila melanogaster.Dev Neurobiol. 2015 Feb;75(2):109-30. doi: 10.1002/dneu.22221. Epub 2014 Aug 5. Dev Neurobiol. 2015. PMID: 25081222 Review.
Cited by
-
Somatotopic organization among parallel sensory pathways that promote a grooming sequence in Drosophila.bioRxiv [Preprint]. 2023 Dec 15:2023.02.11.528119. doi: 10.1101/2023.02.11.528119. bioRxiv. 2023. Update in: Elife. 2024 Apr 18;12:RP87602. doi: 10.7554/eLife.87602. PMID: 36798384 Free PMC article. Updated. Preprint.
-
Male-male interactions shape mate selection in Drosophila.bioRxiv [Preprint]. 2023 Nov 5:2023.11.03.565582. doi: 10.1101/2023.11.03.565582. bioRxiv. 2023. Update in: Cell. 2025 Mar 20;188(6):1486-1503.e25. doi: 10.1016/j.cell.2025.01.008. PMID: 37961193 Free PMC article. Updated. Preprint.
-
Divergent neural circuits for proprioceptive and exteroceptive sensing of the Drosophila leg.Nat Commun. 2025 May 2;16(1):4105. doi: 10.1038/s41467-025-59302-3. Nat Commun. 2025. PMID: 40316553 Free PMC article.
-
Central projections from Johnston's organ in the locust: Axogenesis and brain neuroarchitecture.Dev Genes Evol. 2023 Dec;233(2):147-159. doi: 10.1007/s00427-023-00710-0. Epub 2023 Sep 11. Dev Genes Evol. 2023. PMID: 37695323 Free PMC article.
-
Stereotyped Combination of Hearing and Wind/Gravity-Sensing Neurons in the Johnston's Organ of Drosophila.Front Physiol. 2020 Jan 8;10:1552. doi: 10.3389/fphys.2019.01552. eCollection 2019. Front Physiol. 2020. PMID: 31969834 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
