

EMC10 governs male fertility via maintaining sperm ion balance
- PMID: 29659949
- PMCID: PMC7962889
- DOI: 10.1093/jmcb/mjy024
EMC10 governs male fertility via maintaining sperm ion balance
Abstract
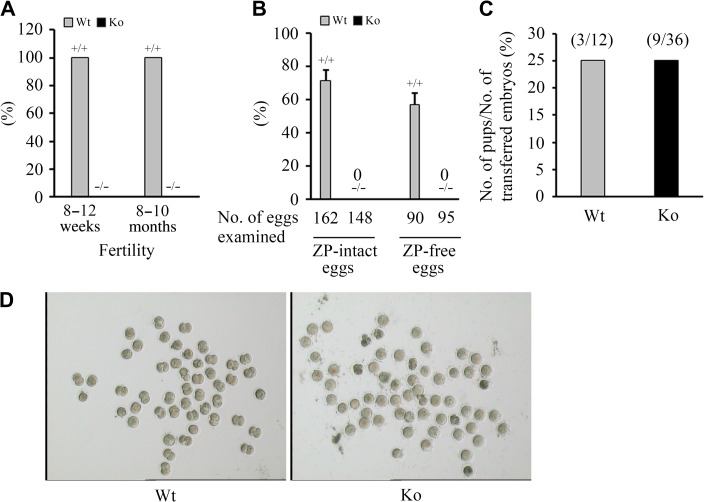
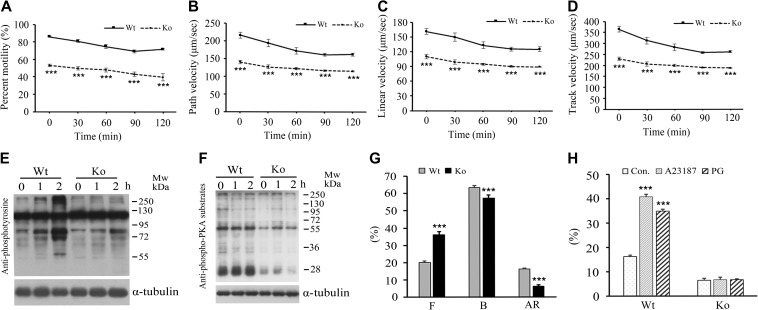
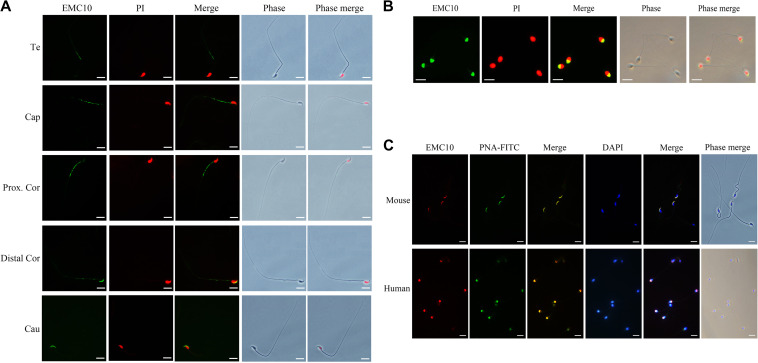
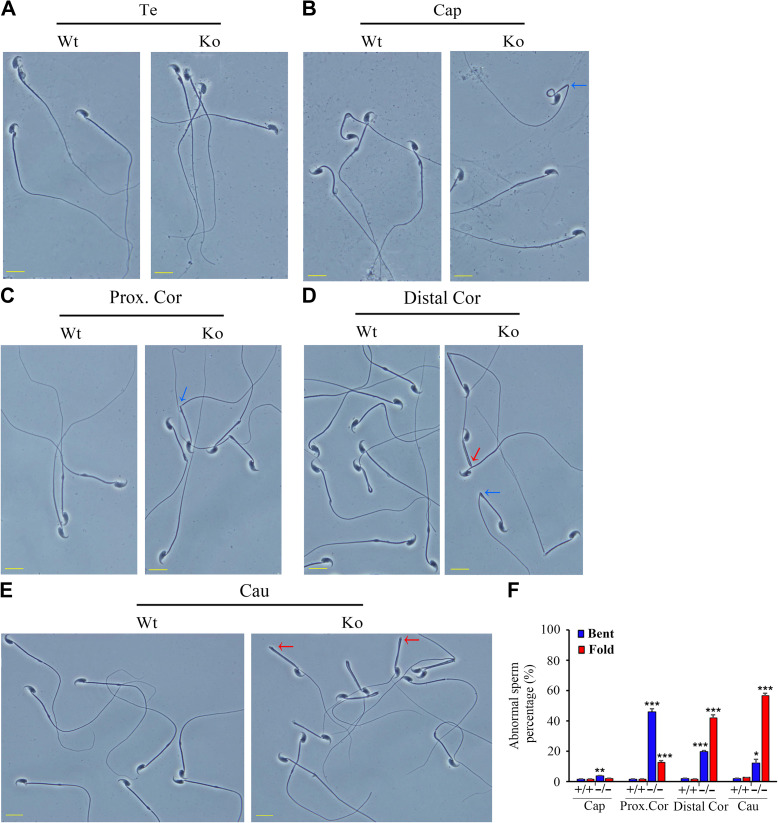
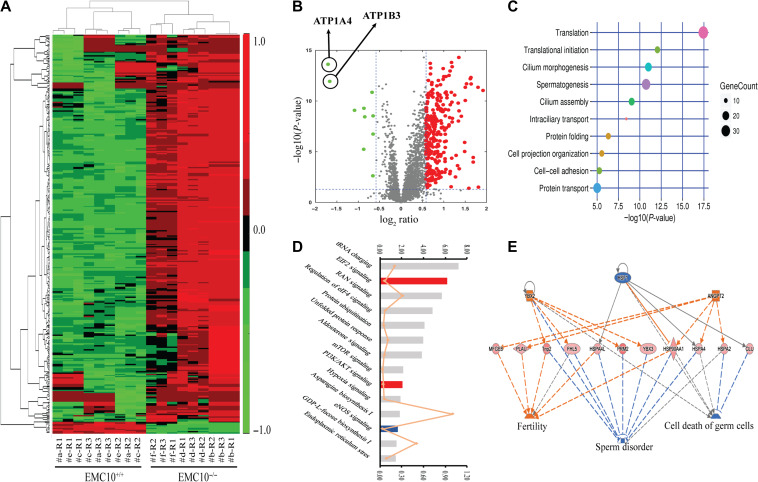
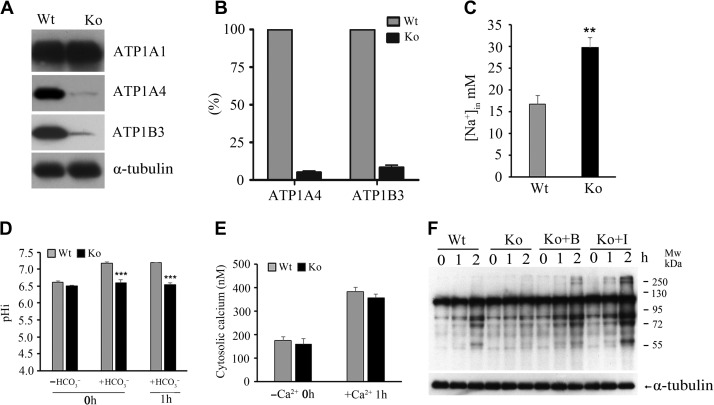
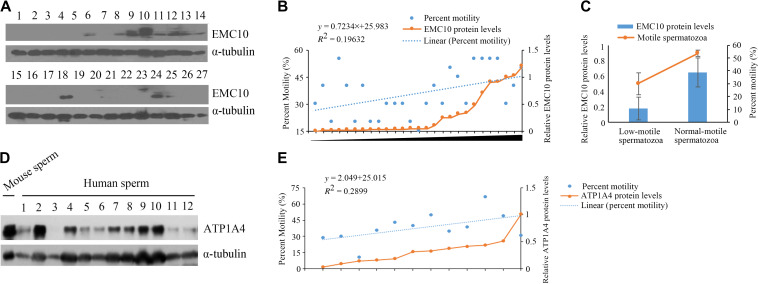
Infertility is a severe public health problem worldwide that prevails up to 15% in reproductive-age couples, and male infertility accounts for half of total infertility. Studies on genetically modified animal models have identified lots of genes involved in the pathogenesis of male infertility. The underlying causes, however, remain largely unclear. In this study, we provide evidence that EMC10, one subunit of endoplasmic reticulum (ER) membrane protein complex (EMC), is required for male fertility. EMC10 is significantly decreased in spermatozoa from patients with asthenozoospermia and positively associated with human sperm motility. Male mice lacking Emc10 gene are completely sterile. Emc10-null spermatozoa exhibit multiple defects including abnormal morphology, decreased motility, impaired capacitation, and impotency of acrosome reaction, thereby which are incapable of fertilizing intact or ZP-free oocytes. However, intracytoplasmic sperm injection could rescue this defect caused by EMC10 deletion. Mechanistically, EMC10 deficiency leads to inactivation of Na/K-ATPase, in turn giving rise to an increased level of intracellular Na+ in spermatozoa, which contributes to decreased sperm motility and abnormal morphology. Other mechanistic investigations demonstrate that the absence of EMC10 results in a reduction of HCO3- entry and subsequent decreases of both cAMP-dependent protein kinase A substrate phosphorylation and protein tyrosine phosphorylation. These data demonstrate that EMC10 is indispensable to male fertility via maintaining sperm ion balance of Na+ and HCO3-, and also suggest that EMC10 is a promising biomarker for male fertility and a potential pharmaceutical target to treat male infertility.
Figures
References
-
- Abou-haila, A., and Tulsiani, D.R. (2009). Signal transduction pathways that regulate sperm capacitation and the acrosome reaction. Arch. Biochem. Biophys. 485, 72–81. - PubMed
-
- Baker, M.A., Hetherington, L., and Aitken, R.J. (2006). Identification of SRC as a key PKA-stimulated tyrosine kinase involved in the capacitation-associated hyperactivation of murine spermatozoa. J. Cell Sci. 119, 3182–3192. - PubMed
-
- Cooper, T.G., Yeung, C.H., Wagenfeld, A., et al. . (2004). Mouse models of infertility due to swollen spermatozoa. Mol. Cell. Endocrinol. 216, 55–63. - PubMed
-
- Darszon, A., Acevedo, J.J., Galindo, B.E., et al. . (2006). Sperm channel diversity and functional multiplicity. Reproduction 131, 977–988. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
