

Historical perspective: phosphatidylserine and phosphatidylethanolamine from the 1800s to the present
- PMID: 29661786
- PMCID: PMC5983396
- DOI: 10.1194/jlr.R084004
Historical perspective: phosphatidylserine and phosphatidylethanolamine from the 1800s to the present
Abstract
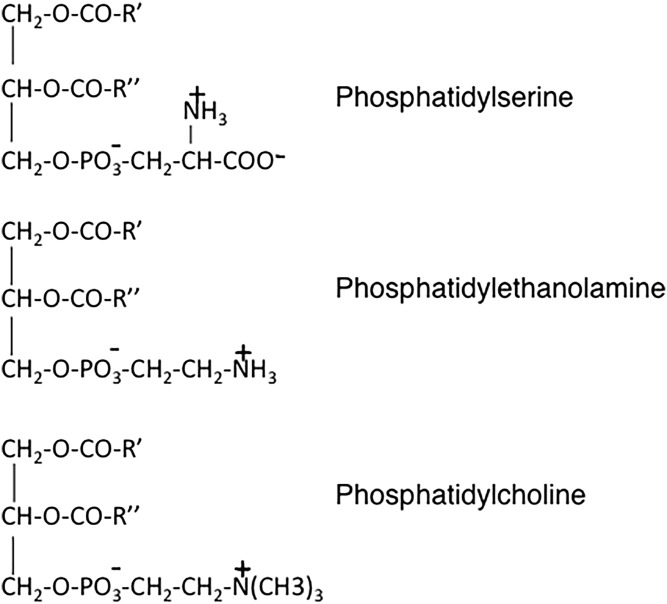
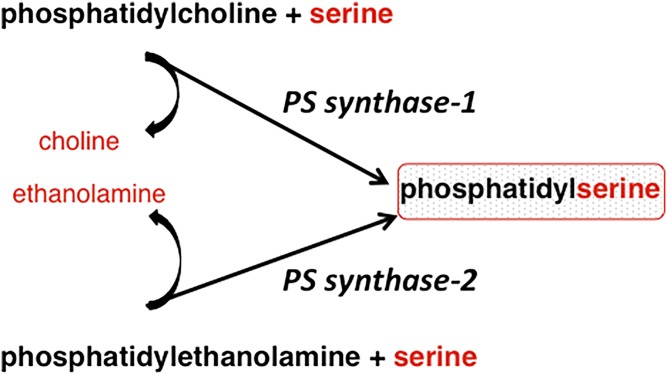
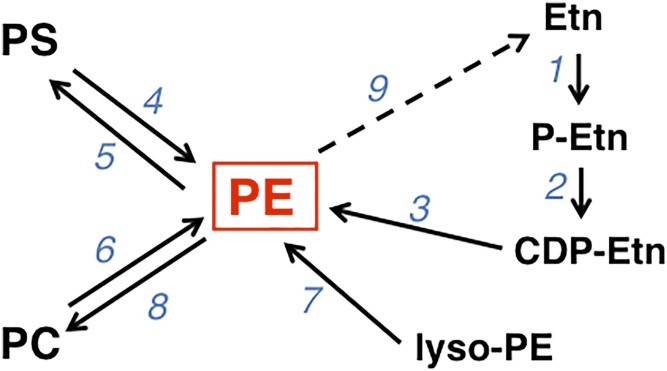
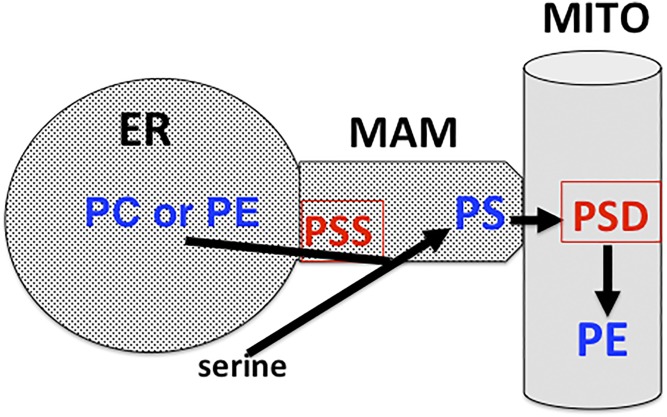
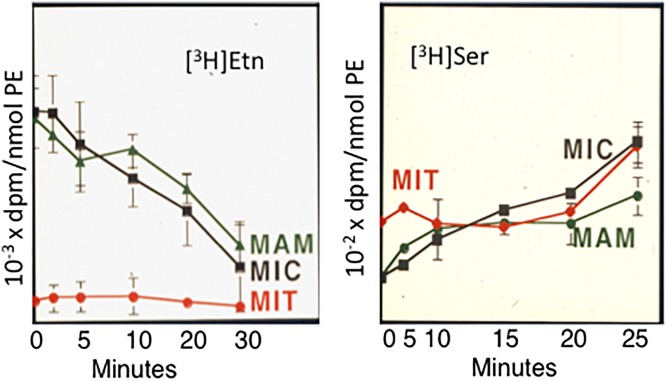
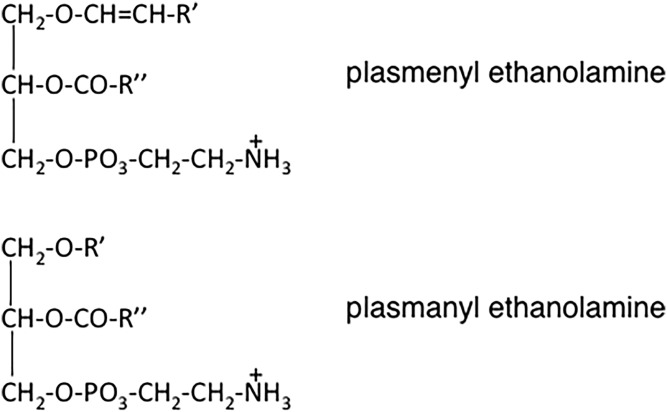
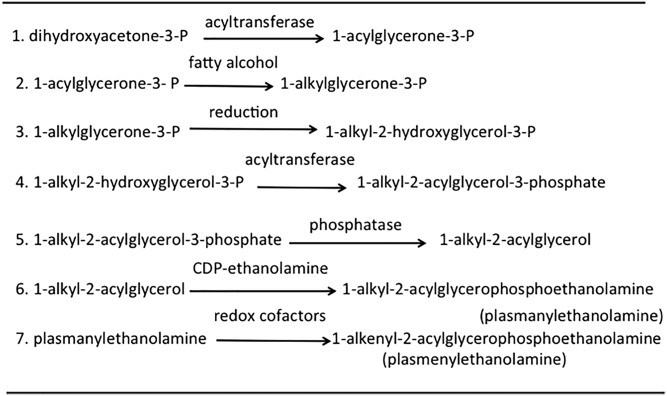
This article provides a historical account of the discovery, chemistry, and biochemistry of two ubiquitous phosphoglycerolipids, phosphatidylserine (PS) and phosphatidylethanolamine (PE), including the ether lipids. In addition, the article describes the biosynthetic pathways for these phospholipids and how these pathways were elucidated. Several unique functions of PS and PE in mammalian cells in addition to their ability to define physical properties of membranes are discussed. For example, the translocation of PS from the inner to the outer leaflet of the plasma membrane of cells occurs during apoptosis and during some other specific physiological processes, and this translocation is responsible for profound life-or-death events. Moreover, mitochondrial function is severely impaired when the PE content of mitochondria is reduced below a threshold level. The discovery and implications of the existence of membrane contact sites between the endoplasmic reticulum and mitochondria and their relevance for PS and PE metabolism, as well as for mitochondrial function, are also discussed. Many of the recent advances in these fields are due to the use of isotope labeling for tracing biochemical pathways. In addition, techniques for disruption of specific genes in mice are now widely used and have provided major breakthroughs in understanding the roles and metabolism of PS and PE in vivo.
Keywords: ether lipids; membranes; mitochondria; phosphoglycerolipids; phospholipid trafficking.
Copyright © 2018 Vance.
Figures


References
-
- Danielli J. F., and Davson H.. 1935. A contribution to the theory of permeability of thin films. J. Cell. Comp. Physiol. 5: 495–547.
-
- Singer S. J., and Nicolson G. L.. 1972. The fluid mosaic model of the structure of cell membranes. Science. 175: 720–731. - PubMed
-
- Simons K., and Ikonen E.. 1997. Functional rafts in cell membranes. Nature. 387: 569–572. - PubMed
-
- Berridge M. J., and Irvine R. F.. 1984. Inositol trisphosphate, a novel second messenger in cellular signal transduction. Nature. 312: 315–321. - PubMed
-
- Nishizuka Y. 1992. Intracellular signaling by hydrolysis of phospholipids and activation of protein kinase C. Science. 258: 607–614. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
