

HadD, a novel fatty acid synthase type II protein, is essential for alpha- and epoxy-mycolic acid biosynthesis and mycobacterial fitness
- PMID: 29662082
- PMCID: PMC5902629
- DOI: 10.1038/s41598-018-24380-5
HadD, a novel fatty acid synthase type II protein, is essential for alpha- and epoxy-mycolic acid biosynthesis and mycobacterial fitness
Abstract
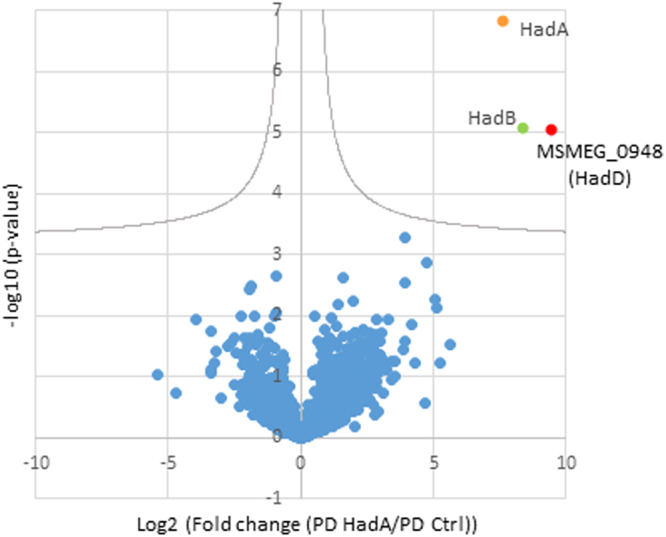
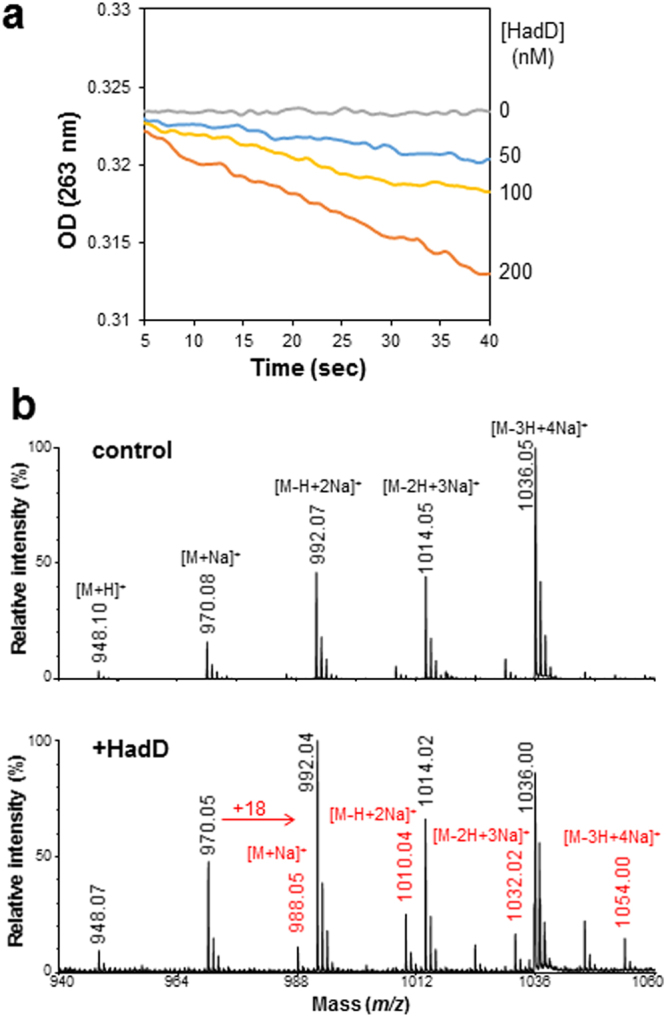
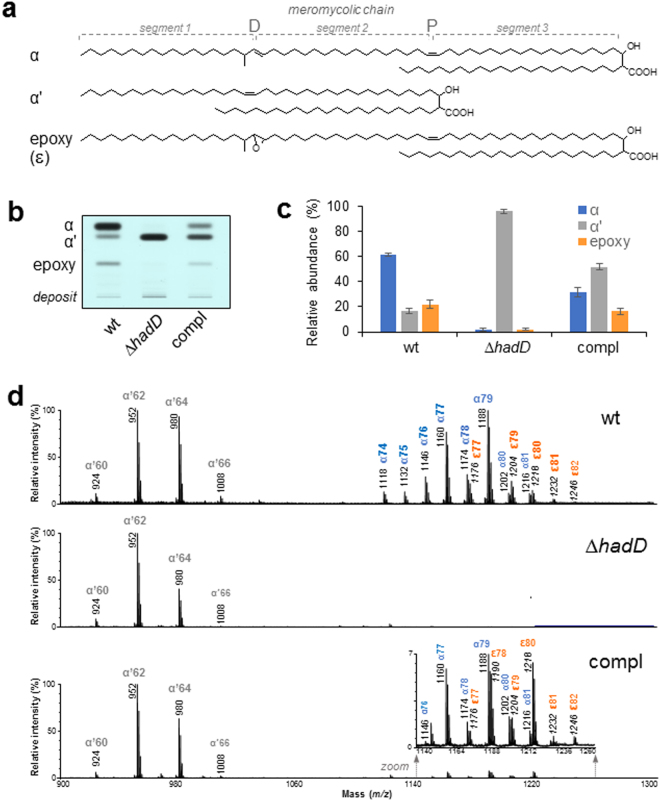
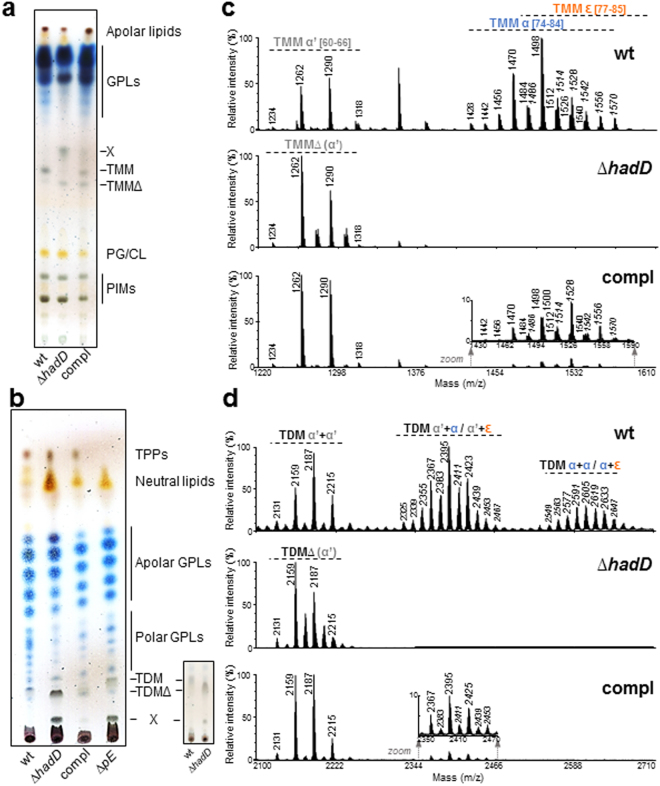
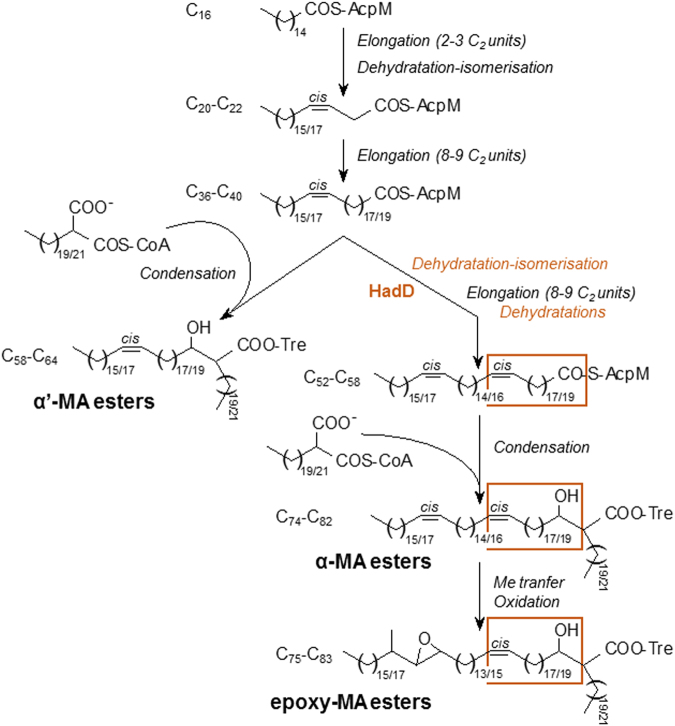
Mycolic acids (MAs) have a strategic location within the mycobacterial envelope, deeply influencing its architecture and permeability, and play a determinant role in the pathogenicity of mycobacteria. The fatty acid synthase type II (FAS-II) multienzyme system is involved in their biosynthesis. A combination of pull-downs and proteomics analyses led to the discovery of a mycobacterial protein, HadD, displaying highly specific interactions with the dehydratase HadAB of FAS-II. In vitro activity assays and homology modeling showed that HadD is, like HadAB, a hot dog folded (R)-specific hydratase/dehydratase. A hadD knockout mutant of Mycobacterium smegmatis produced only the medium-size alpha'-MAs. Data strongly suggest that HadD is involved in building the third meromycolic segment during the late FAS-II elongation cycles, leading to the synthesis of the full-size alpha- and epoxy-MAs. The change in the envelope composition induced by hadD inactivation strongly altered the bacterial fitness and capacities to aggregate, assemble into colonies or biofilms and spread by sliding motility, and conferred a hypersensitivity to the firstline antimycobacterial drug rifampicin. This showed that the cell surface properties and the envelope integrity were greatly affected. With the alarmingly increasing case number of nontuberculous mycobacterial diseases, HadD appears as an attractive target for drug development.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- WHO. Global tuberculosis report 2017. (ed. WHO Press) (World Health Organization, 2017).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
