

Stimulation of entorhinal cortex-dentate gyrus circuitry is antidepressive
- PMID: 29662202
- PMCID: PMC5948139
- DOI: 10.1038/s41591-018-0002-1
Stimulation of entorhinal cortex-dentate gyrus circuitry is antidepressive
Erratum in
-
Publisher Correction: Stimulation of entorhinal cortex-dentate gyrus circuitry is antidepressive.Nat Med. 2018 Sep;24(9):1482. doi: 10.1038/s41591-018-0084-9. Nat Med. 2018. PMID: 29934536
Abstract
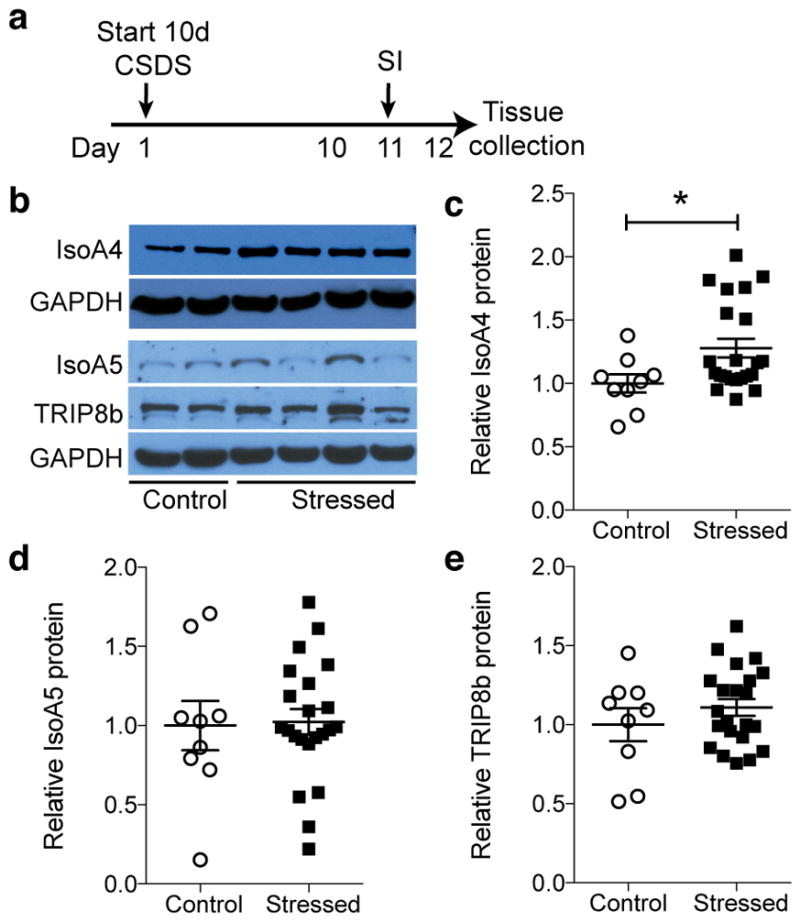
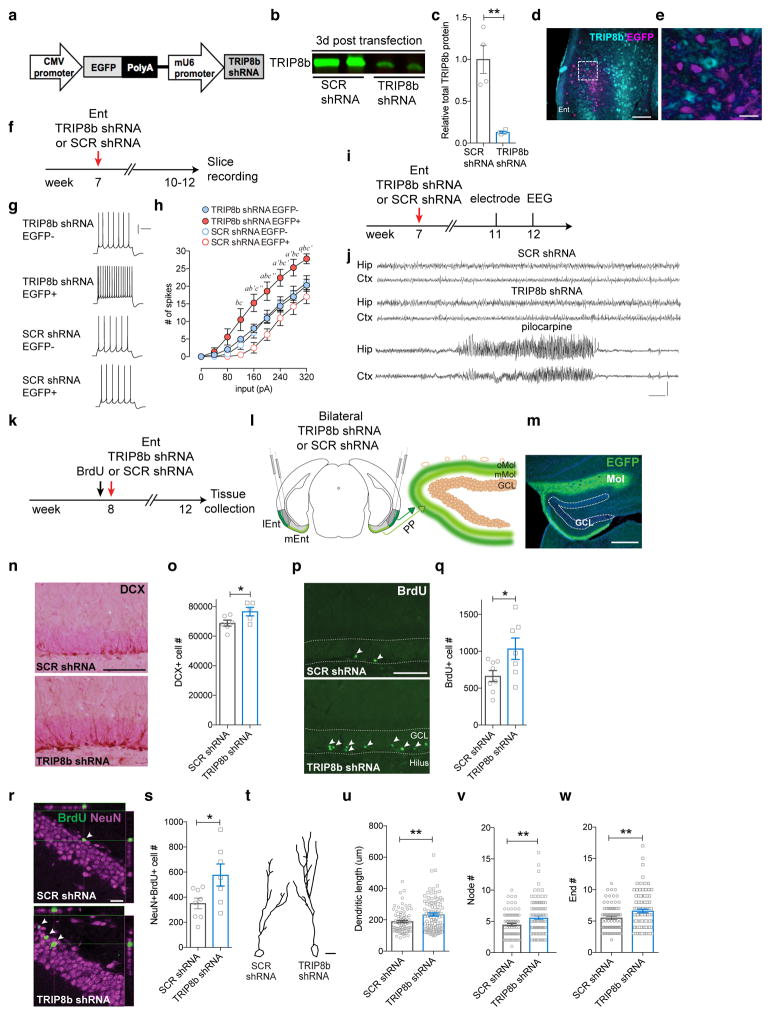
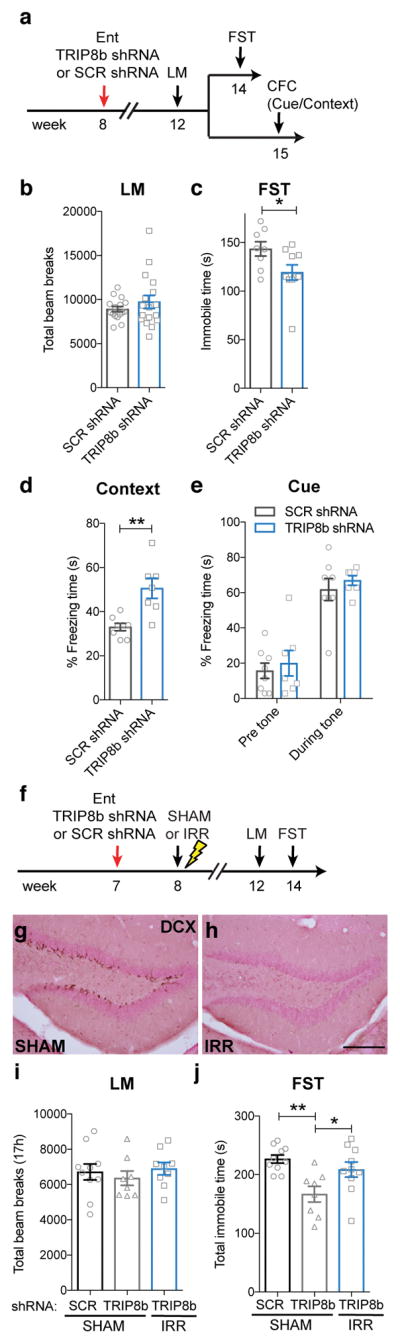
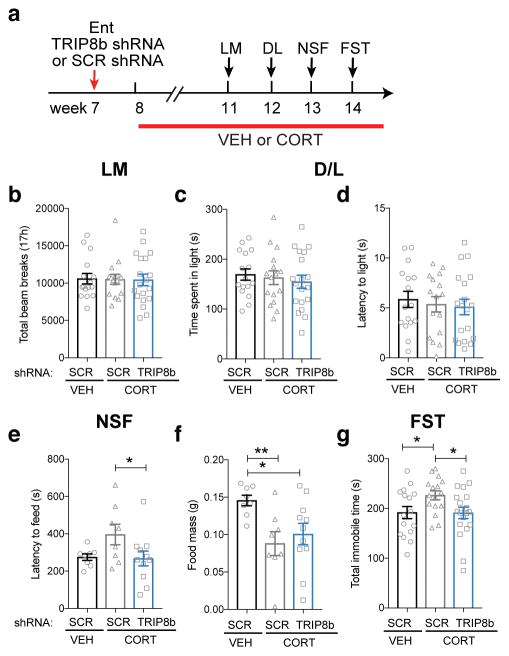
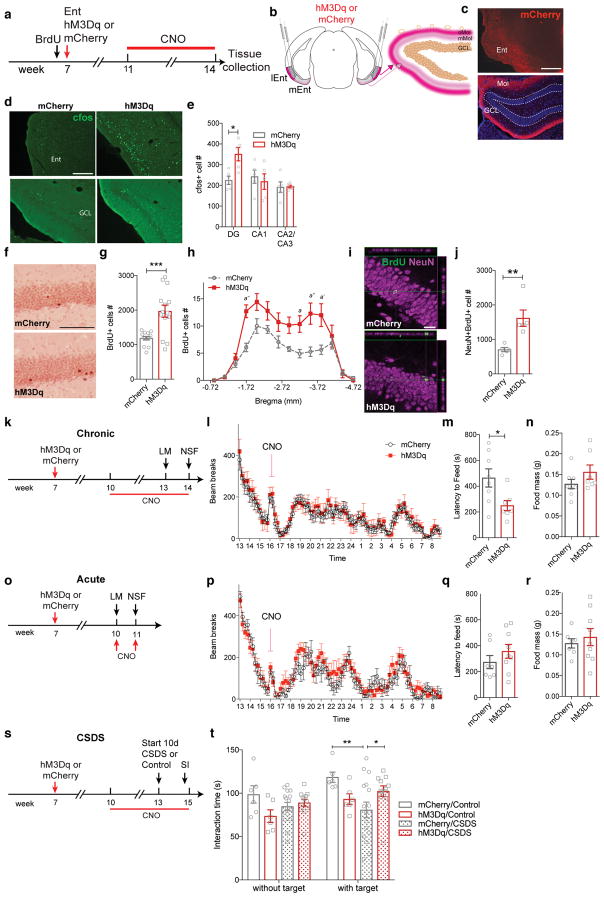
Major depressive disorder (MDD) is considered a 'circuitopathy', and brain stimulation therapies hold promise for ameliorating MDD symptoms, including hippocampal dysfunction. It is unknown whether stimulation of upstream hippocampal circuitry, such as the entorhinal cortex (Ent), is antidepressive, although Ent stimulation improves learning and memory in mice and humans. Here we show that molecular targeting (Ent-specific knockdown of a psychosocial stress-induced protein) and chemogenetic stimulation of Ent neurons induce antidepressive-like effects in mice. Mechanistically, we show that Ent-stimulation-induced antidepressive-like behavior relies on the generation of new hippocampal neurons. Thus, controlled stimulation of Ent hippocampal afferents is antidepressive via increased hippocampal neurogenesis. These findings emphasize the power and potential of Ent glutamatergic afferent stimulation-previously well-known for its ability to influence learning and memory-for MDD treatment.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
