

Myelination of the developing lateral olfactory tract and anterior commissure
- PMID: 29665005
- PMCID: PMC5990462
- DOI: 10.1002/cne.24452
Myelination of the developing lateral olfactory tract and anterior commissure
Abstract
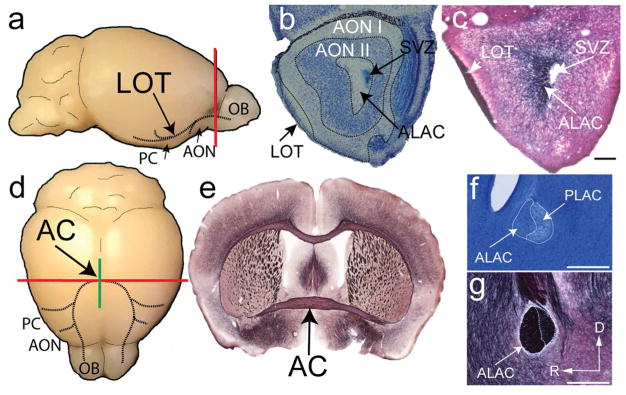
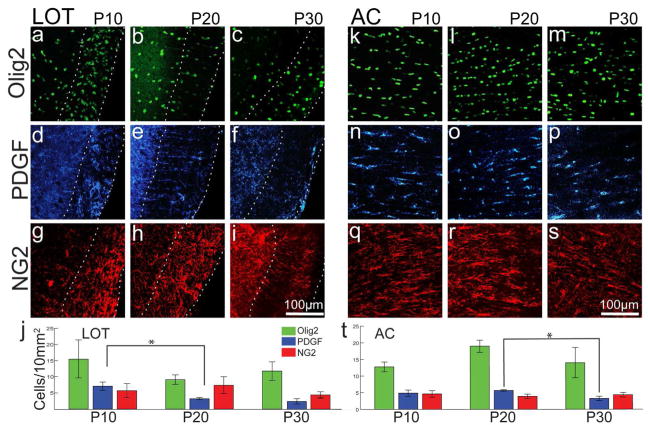
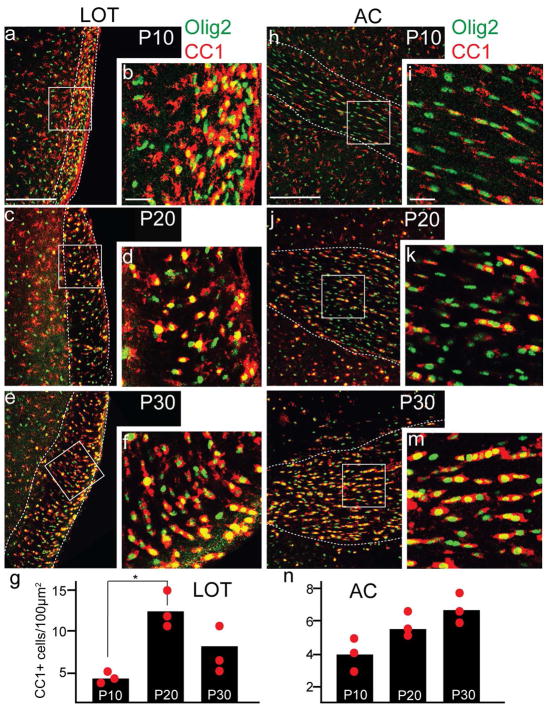
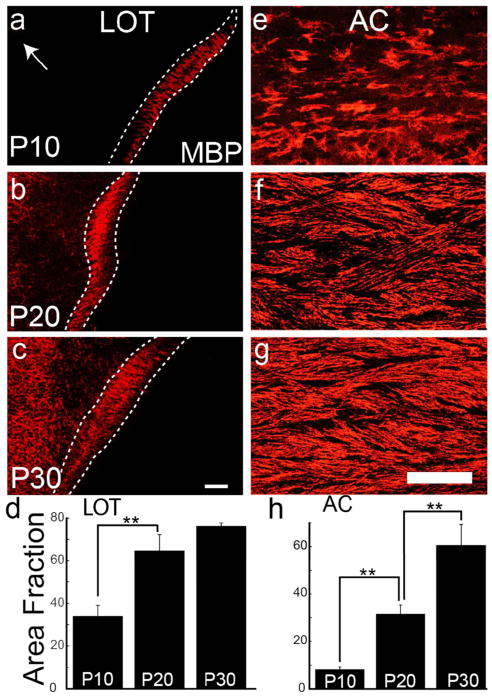
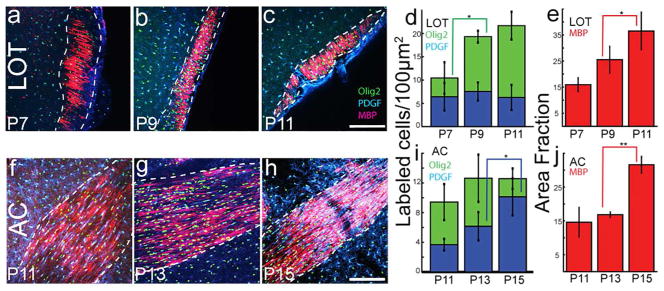
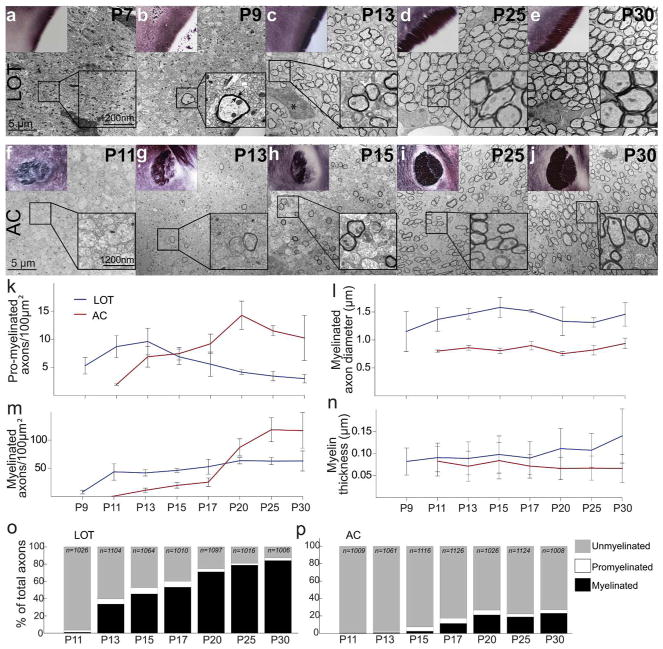
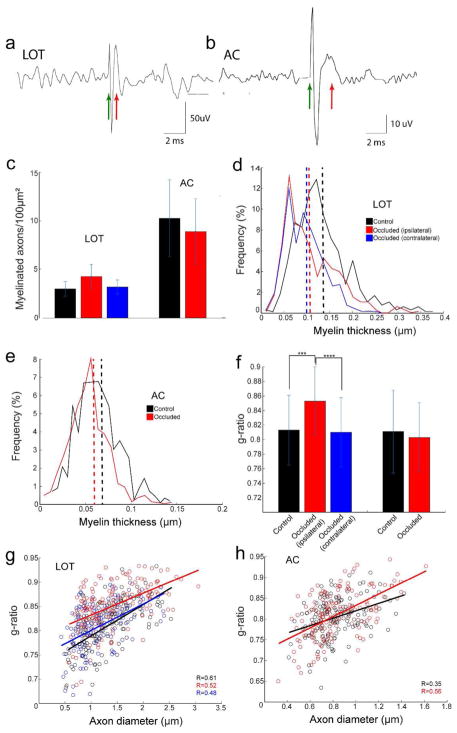
Both the lateral olfactory tract (LOT) and anterior limb of the anterior commissure (AC) carry olfactory information. The LOT forms the projection from the olfactory bulb to the ipsilateral olfactory cortices, while the AC carries odor information across the midline to the contralateral olfactory cortex and bulb. The LOT and AC differ on a number of dimensions, including early development and functional onset. The present work, examining their myelination in mice, reveals additional important differences. For example, the LOT initiates myelination 3-4 days earlier than the AC, evidenced by both an earlier increase in myelin basic protein staining seen with immunohistochemistry and an earlier appearance of myelinated fibers using electron microscopy. While both exhibit a period of rapid myelination, it occurs 4-5 days earlier in the LOT than the AC. The tracts also respond differently to early sensory restriction. Unilateral naris occlusion from the day after birth to postnatal day 30 had no consistent effects on the AC but resulted in significantly thinner myelin sheaths relative to axon caliber in the LOT. Finally, the two tracts differ structurally (the LOT contains larger, more densely packed axons with significantly thicker myelin sheaths resulting in a conduction velocity that is more than twice as fast as the AC). The findings indicate that these two large, accessible tracts provide an important means for studying brain maturation due to basic differences in both the timing of their maturation and general organization.
Keywords: RRID: AB_10141047; RRID: AB_11213678; RRID: AB_2057371; RRID: AB_2140491; RRID: AB_2236897; electron microscopy; immunohistochemistry; myelin precursors; olfactory cortex; oligodendrocytes.
© 2018 Wiley Periodicals, Inc.
Conflict of interest statement
This research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Experimental Demyelination of the Lateral Olfactory Tract and Anterior Commissure.Neuroscience. 2020 May 10;434:93-101. doi: 10.1016/j.neuroscience.2020.03.026. Epub 2020 Mar 26. Neuroscience. 2020. PMID: 32224229
-
Acetyl-l-carnitine enhances myelination of regenerated fibers of the lateral olfactory tract.Neurosci Lett. 2017 Jul 13;653:215-219. doi: 10.1016/j.neulet.2017.06.001. Epub 2017 Jun 3. Neurosci Lett. 2017. PMID: 28583580
-
Excitable Axonal Domains Adapt to Sensory Deprivation in the Olfactory System.J Neurosci. 2022 Feb 23;42(8):1491-1509. doi: 10.1523/JNEUROSCI.0305-21.2021. Epub 2022 Jan 12. J Neurosci. 2022. PMID: 35022219 Free PMC article.
-
Activity-Dependent Myelination.Adv Exp Med Biol. 2019;1190:43-51. doi: 10.1007/978-981-32-9636-7_4. Adv Exp Med Biol. 2019. PMID: 31760637 Review.
-
White matter plasticity in adulthood.Neuroscience. 2014 Sep 12;276:148-60. doi: 10.1016/j.neuroscience.2013.10.018. Epub 2013 Oct 23. Neuroscience. 2014. PMID: 24161723 Review.
Cited by
-
Ambient sound stimulation tunes axonal conduction velocity by regulating radial growth of myelin on an individual, axon-by-axon basis.Proc Natl Acad Sci U S A. 2024 Mar 12;121(11):e2316439121. doi: 10.1073/pnas.2316439121. Epub 2024 Mar 5. Proc Natl Acad Sci U S A. 2024. PMID: 38442165 Free PMC article.
-
The mouse olfactory peduncle 4: Development of synapses, perineuronal nets, and capillaries.J Comp Neurol. 2020 Mar 1;528(4):637-649. doi: 10.1002/cne.24778. Epub 2019 Oct 18. J Comp Neurol. 2020. PMID: 31571216 Free PMC article.
-
Development of piriform cortex interhemispheric connections via the anterior commissure: progressive and regressive strategies.Brain Struct Funct. 2018 Dec;223(9):4067-4085. doi: 10.1007/s00429-018-1741-y. Epub 2018 Aug 24. Brain Struct Funct. 2018. PMID: 30141078 Free PMC article.
-
Embryonic and postnatal development of mouse olfactory tubercle.Mol Cell Neurosci. 2019 Jul;98:82-96. doi: 10.1016/j.mcn.2019.06.002. Epub 2019 Jun 11. Mol Cell Neurosci. 2019. PMID: 31200100 Free PMC article.
-
From Circuits to Lifespan: Translating Mouse and Human Timelines with Neuroimaging-Based Tractography.J Neurosci. 2025 Mar 19;45(12):e1429242025. doi: 10.1523/JNEUROSCI.1429-24.2025. J Neurosci. 2025. PMID: 39870528
References
-
- Aggarwal S, Yurlova L, Simons M. Central nervous system myelin: structure, synthesis, and assembly. Trends in Cell Biology. 2011;21(10):585–593. - PubMed
-
- Asou H, Hamada K, Miyazaki T, Sakota T, Hayashi K, Takeda Y, Marret S, Delpech B, Itoh K, Uyemura K. CNS myelinogenesis in vitro: time course and pattern of rat oligodendrocyte development. Journal of Neuroscience Research. 1995;40(4):519–534. - PubMed
-
- Baumann N, Pham-Dinh D. Biology of oligodendrocyte and myelin in the mammalian central nervous system. Physiological Reviews. 2001;81(2):871–927. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
