

Lmo1656 is a secreted virulence factor of Listeria monocytogenes that interacts with the sorting nexin 6-BAR complex
- PMID: 29666193
- PMCID: PMC6005434
- DOI: 10.1074/jbc.RA117.000365
Lmo1656 is a secreted virulence factor of Listeria monocytogenes that interacts with the sorting nexin 6-BAR complex
Abstract
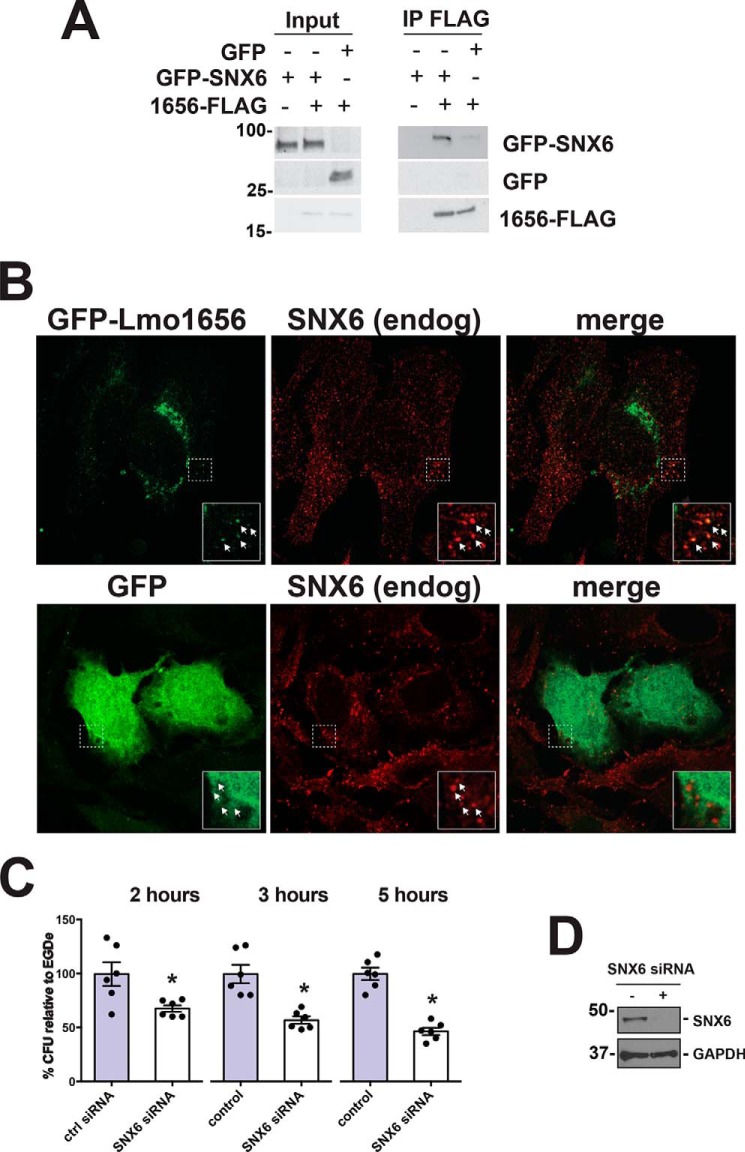
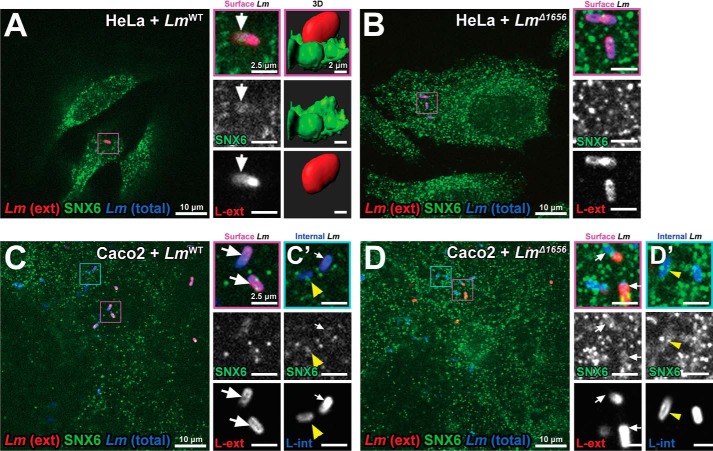
Listeria monocytogenes (Lm) is a facultative intracellular bacterial pathogen and the causative agent of listeriosis, a rare but fatal disease. During infection, Lm can traverse several physiological barriers; it can cross the intestine and placenta barrier and, in immunocompromised individuals, the blood-brain barrier. With the recent plethora of sequenced genomes available for Lm, it is clear that the complete repertoire of genes used by Lm to interact with its host remains to be fully explored. Recently, we focused on secreted Lm proteins because they are likely to interact with host cell components. Here, we investigated a putatively secreted protein of Lm, Lmo1656, that is present in most sequenced strains of Lm but absent in the nonpathogenic species Listeria innocua. lmo1656 gene is predicted to encode a small, positively charged protein. We show that Lmo1656 is secreted by Lm Furthermore, deletion of the lmo1656 gene (Δlmo1656) attenuates virulence in mice infected orally but not intravenously, suggesting that Lmo1656 plays a role during oral listeriosis. We identified sorting nexin 6 (SNX6), an endosomal sorting component and BAR domain-containing protein, as a host cell interactor of Lmol656. SNX6 colocalizes with WT Lm during the early steps of infection. This colocalization depends on Lmo1656, and RNAi of SNX6 impairs infection in infected tissue culture cells, suggesting that SNX6 is utilized by Lm during infection. Our results reveal that Lmo1656 is a novel secreted virulence factor of Lm that facilitates recruitment of a specific member of the sorting nexin family in the mammalian host.
Keywords: Listeria monocytogenes; Lmo1656; bacterial pathogenesis; host-pathogen interaction; infection; sorting nexin (SNX); sorting nexin 6; virulence factor.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
