

Drought delays development of the sorghum root microbiome and enriches for monoderm bacteria
- PMID: 29666229
- PMCID: PMC5939072
- DOI: 10.1073/pnas.1717308115
Drought delays development of the sorghum root microbiome and enriches for monoderm bacteria
Erratum in
-
Correction for Xu et al., Drought delays development of the sorghum root microbiome and enriches for monoderm bacteria.Proc Natl Acad Sci U S A. 2018 May 22;115(21):E4952. doi: 10.1073/pnas.1807275115. Epub 2018 May 14. Proc Natl Acad Sci U S A. 2018. PMID: 29760104 Free PMC article. No abstract available.
Abstract
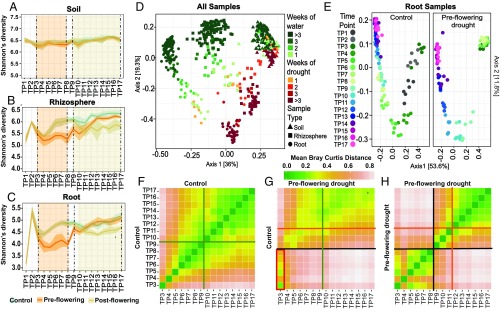
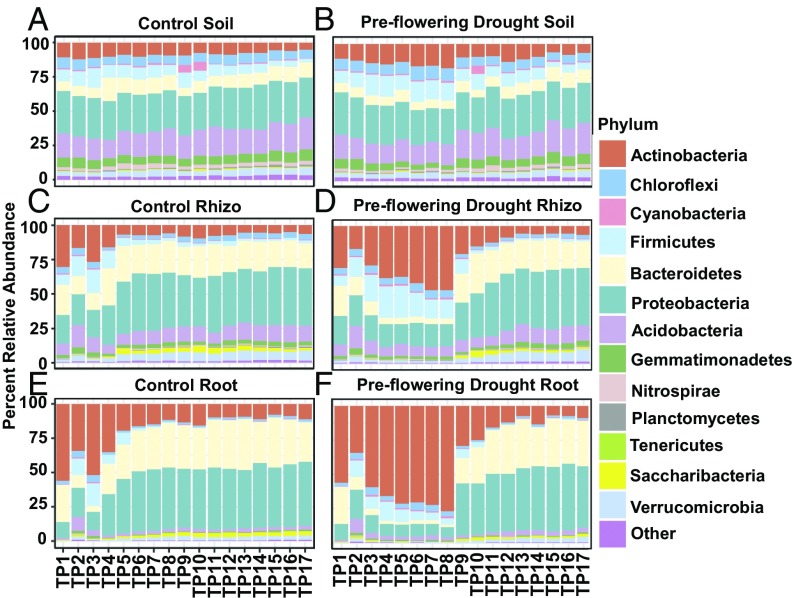
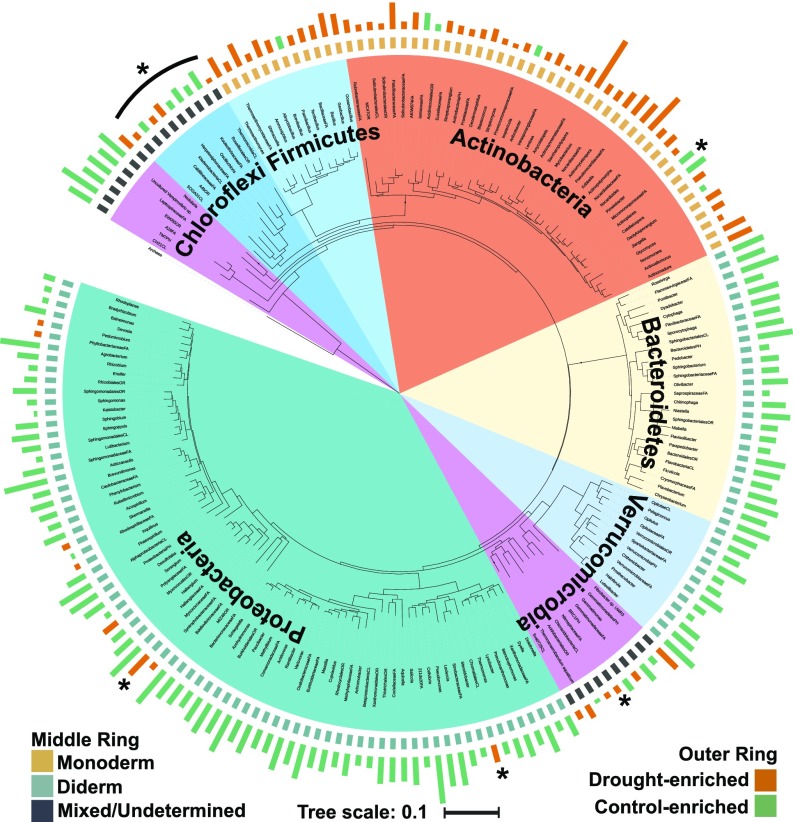
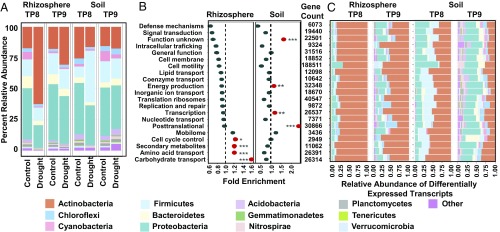
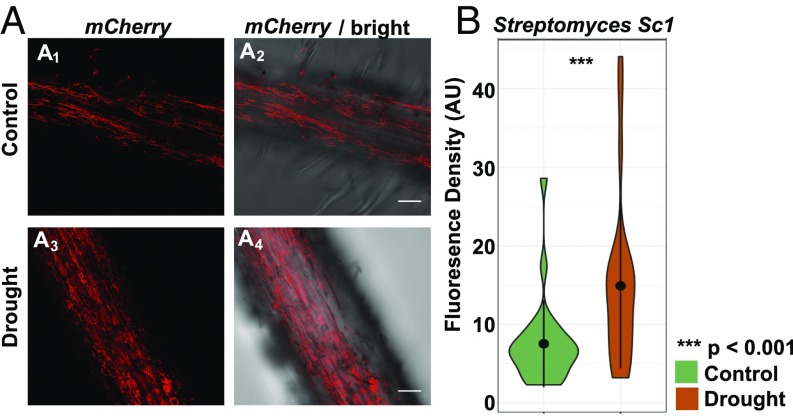
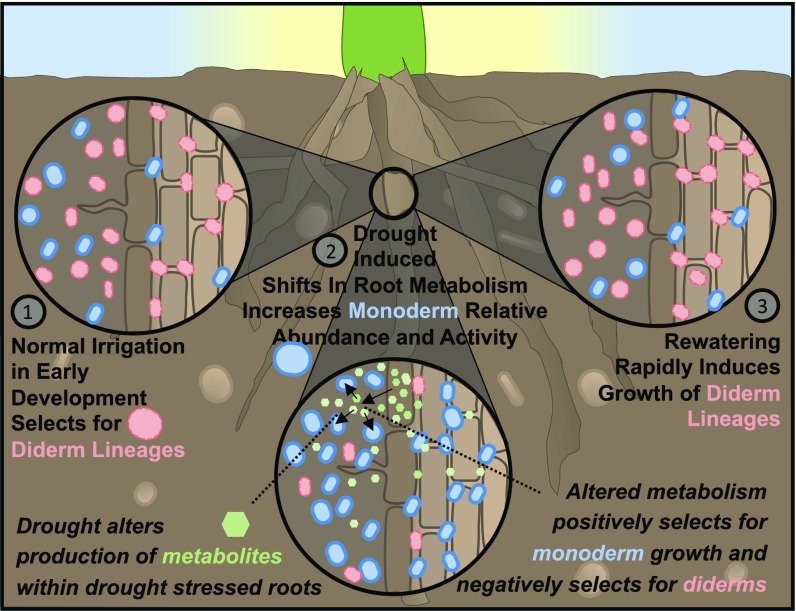
Drought stress is a major obstacle to crop productivity, and the severity and frequency of drought are expected to increase in the coming century. Certain root-associated bacteria have been shown to mitigate the negative effects of drought stress on plant growth, and manipulation of the crop microbiome is an emerging strategy for overcoming drought stress in agricultural systems, yet the effect of drought on the development of the root microbiome is poorly understood. Through 16S rRNA amplicon and metatranscriptome sequencing, as well as root metabolomics, we demonstrate that drought delays the development of the early sorghum root microbiome and causes increased abundance and activity of monoderm bacteria, which lack an outer cell membrane and contain thick cell walls. Our data suggest that altered plant metabolism and increased activity of bacterial ATP-binding cassette (ABC) transporter genes are correlated with these shifts in community composition. Finally, inoculation experiments with monoderm isolates indicate that increased colonization of the root during drought can positively impact plant growth. Collectively, these results demonstrate the role that drought plays in restructuring the root microbiome and highlight the importance of temporal sampling when studying plant-associated microbiomes.
Keywords: drought; metatranscriptome; microbiome; root; sorghum.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
