

Desynchronization of slow oscillations in the basal ganglia during natural sleep
- PMID: 29666271
- PMCID: PMC5939089
- DOI: 10.1073/pnas.1720795115
Desynchronization of slow oscillations in the basal ganglia during natural sleep
Abstract
Slow oscillations of neuronal activity alternating between firing and silence are a hallmark of slow-wave sleep (SWS). These oscillations reflect the default activity present in all mammalian species, and are ubiquitous to anesthesia, brain slice preparations, and neuronal cultures. In all these cases, neuronal firing is highly synchronous within local circuits, suggesting that oscillation-synchronization coupling may be a governing principle of sleep physiology regardless of anatomical connectivity. To investigate whether this principle applies to overall brain organization, we recorded the activity of individual neurons from basal ganglia (BG) structures and the thalamocortical (TC) network over 70 full nights of natural sleep in two vervet monkeys. During SWS, BG neurons manifested slow oscillations (∼0.5 Hz) in firing rate that were as prominent as in the TC network. However, in sharp contrast to any neural substrate explored thus far, the slow oscillations in all BG structures were completely desynchronized between individual neurons. Furthermore, whereas in the TC network single-cell spiking was locked to slow oscillations in the local field potential (LFP), the BG LFP exhibited only weak slow oscillatory activity and failed to entrain nearby cells. We thus show that synchrony is not inherent to slow oscillations, and propose that the BG desynchronization of slow oscillations could stem from its unique anatomy and functional connectivity. Finally, we posit that BG slow-oscillation desynchronization may further the reemergence of slow-oscillation traveling waves from multiple independent origins in the frontal cortex, thus significantly contributing to normal SWS.
Keywords: basal ganglia; desynchronization; nonhuman primate; sleep; slow oscillations.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
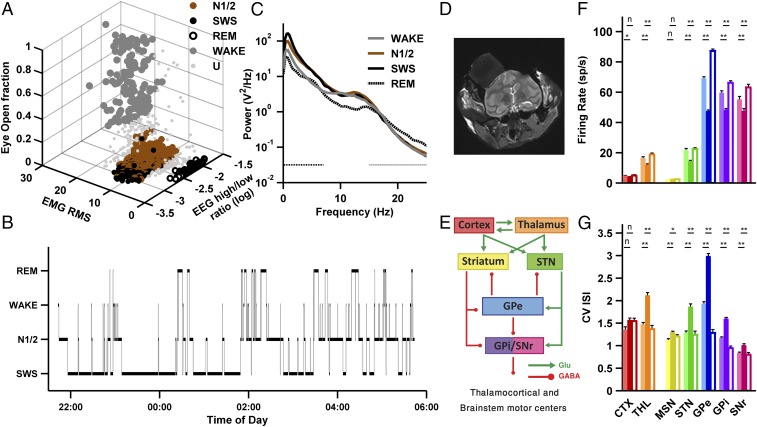
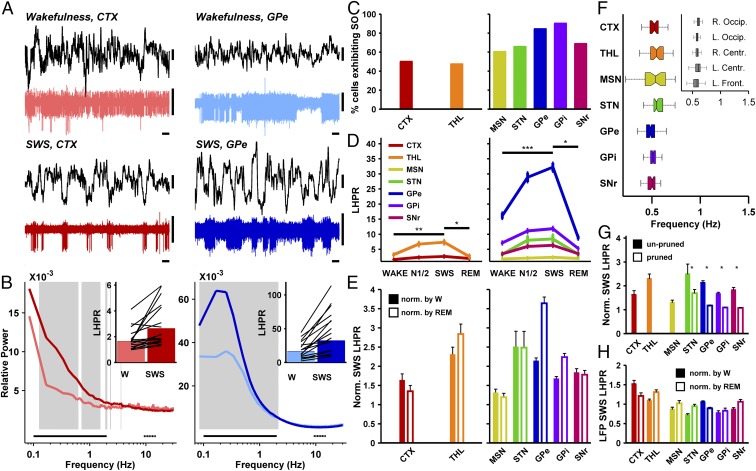
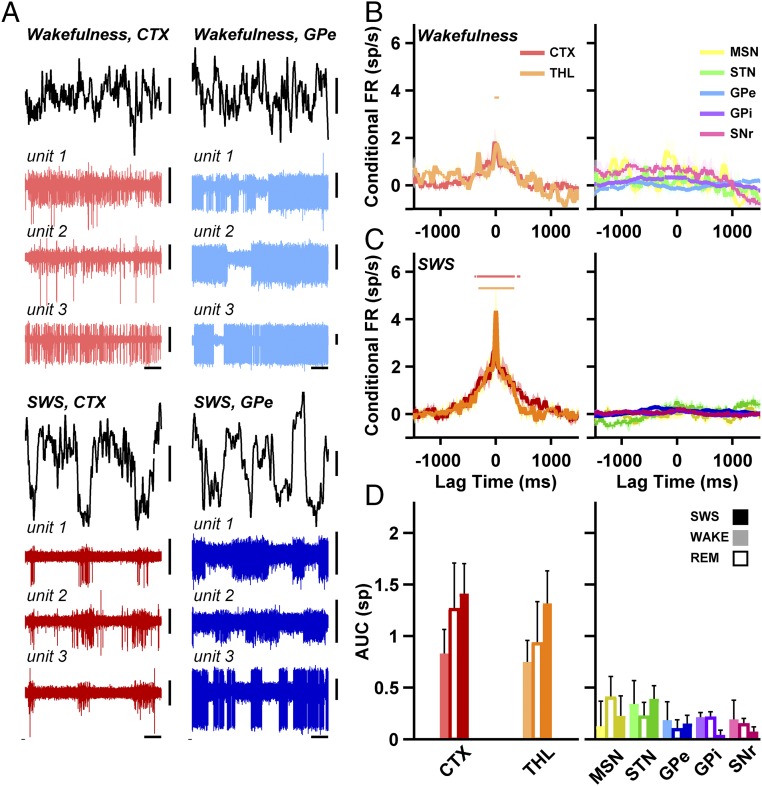
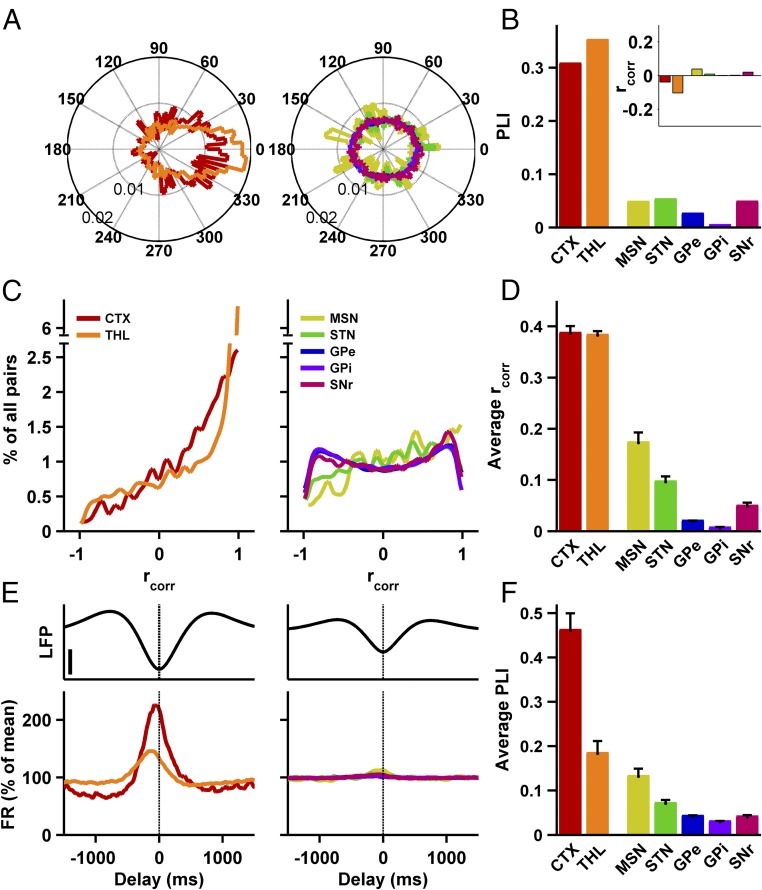
References
-
- Nir Y, Massimini M, Boly M, Tononi G. Sleep and consciousness. In: Cavanna AE, Nani A, Blumenfeld H, Laureys S, editors. Neuroimaging of Consciousness. Springer; Berlin: 2013. pp. 133–182.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
