

Determining the Specificity of Cascade Binding, Interference, and Primed Adaptation In Vivo in the Escherichia coli Type I-E CRISPR-Cas System
- PMID: 29666291
- PMCID: PMC5904413
- DOI: 10.1128/mBio.02100-17
Determining the Specificity of Cascade Binding, Interference, and Primed Adaptation In Vivo in the Escherichia coli Type I-E CRISPR-Cas System
Abstract
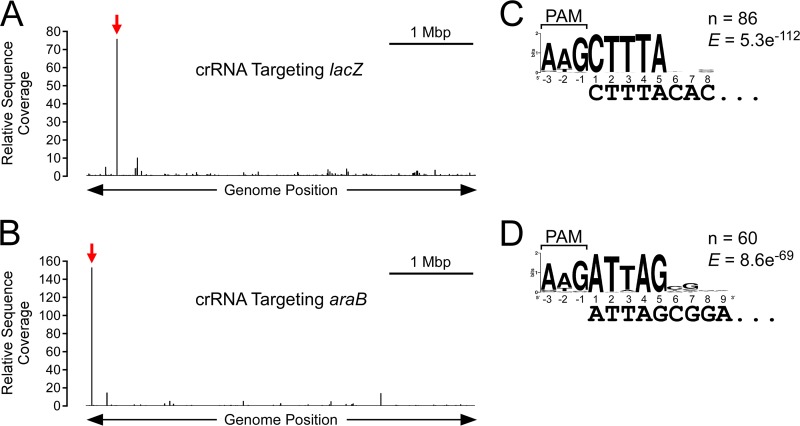
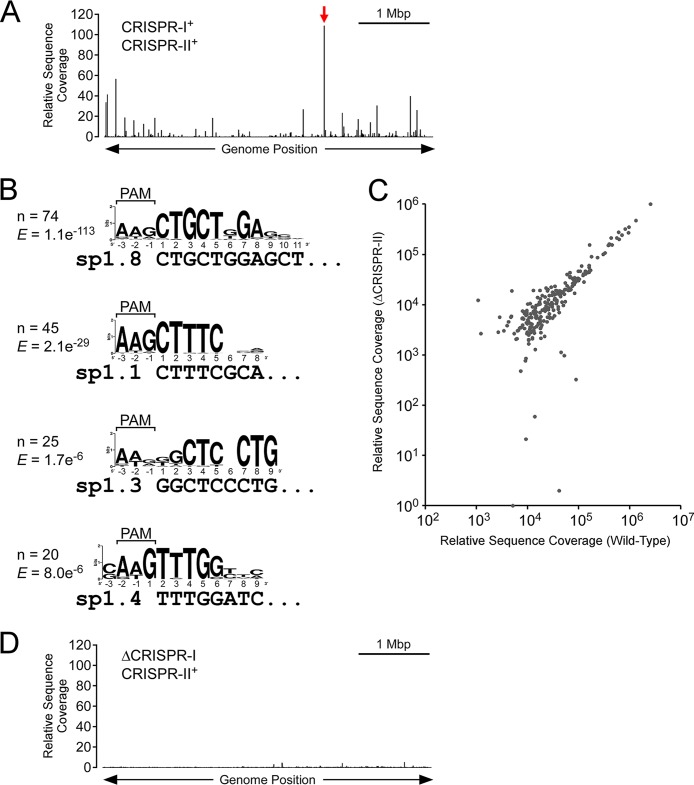
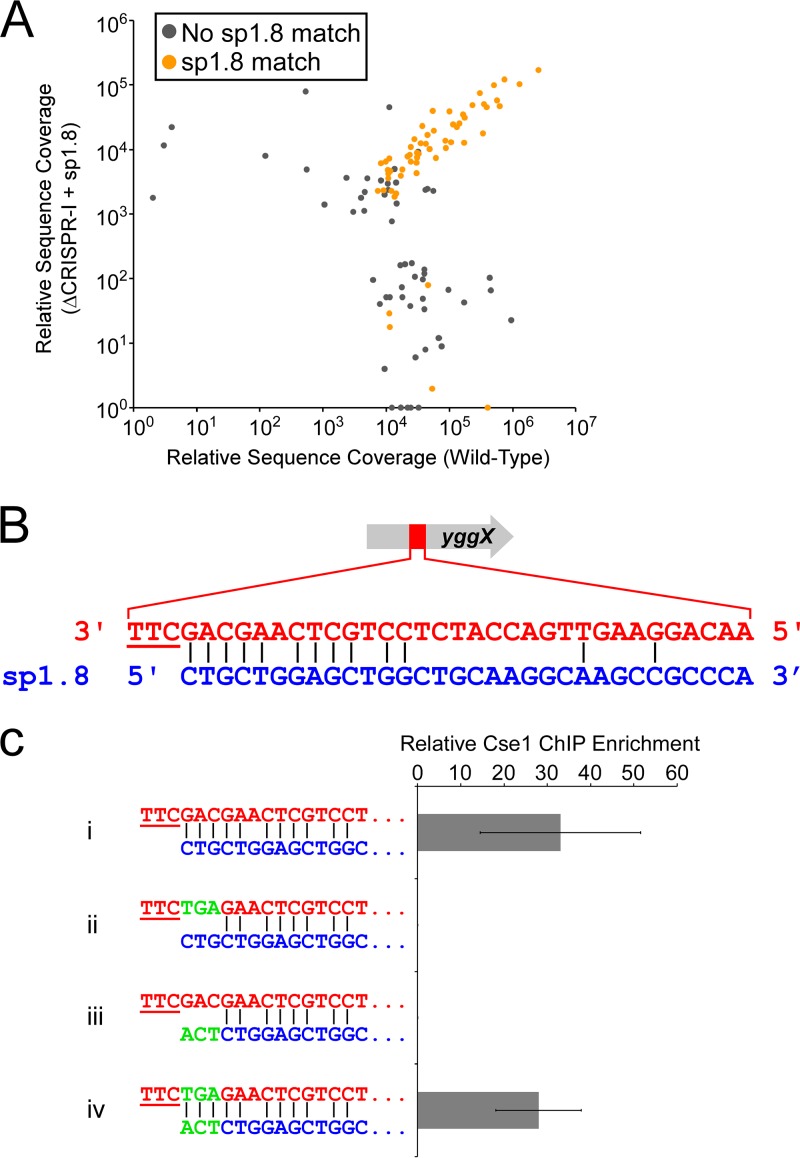
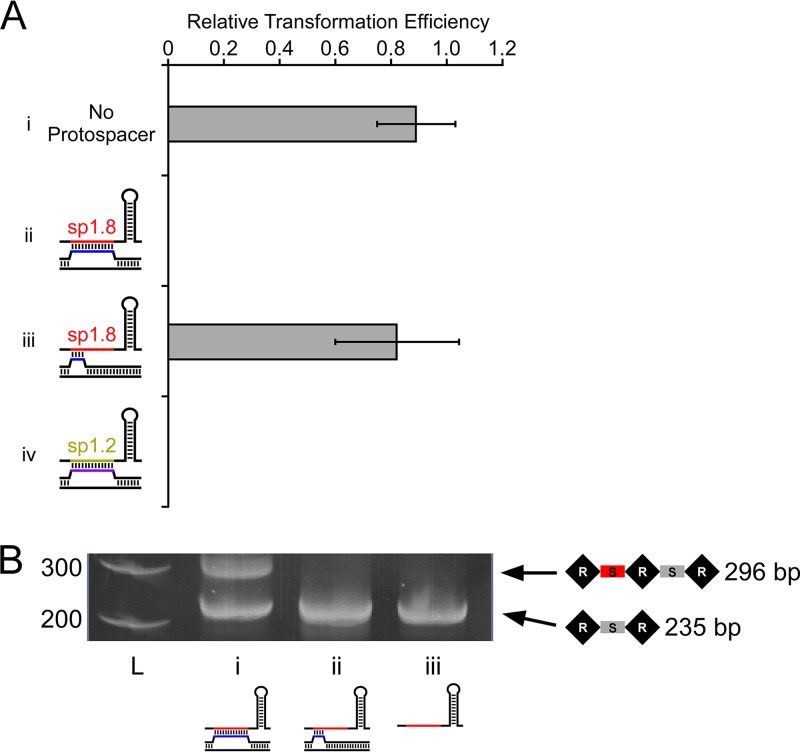
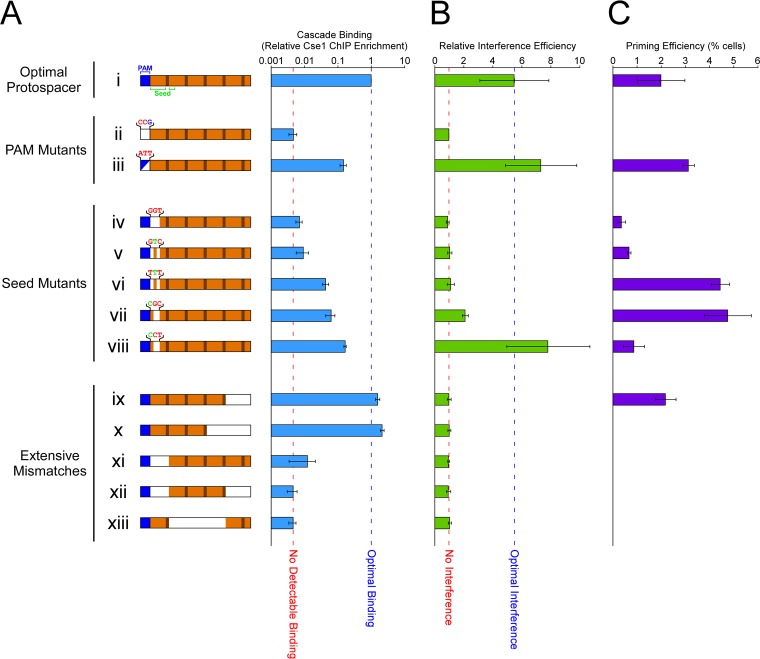
In clustered regularly interspaced short palindromic repeat (CRISPR)-Cas (CRISPR-associated) immunity systems, short CRISPR RNAs (crRNAs) are bound by Cas proteins, and these complexes target invading nucleic acid molecules for degradation in a process known as interference. In type I CRISPR-Cas systems, the Cas protein complex that binds DNA is known as Cascade. Association of Cascade with target DNA can also lead to acquisition of new immunity elements in a process known as primed adaptation. Here, we assess the specificity determinants for Cascade-DNA interaction, interference, and primed adaptation in vivo, for the type I-E system of Escherichia coli Remarkably, as few as 5 bp of crRNA-DNA are sufficient for association of Cascade with a DNA target. Consequently, a single crRNA promotes Cascade association with numerous off-target sites, and the endogenous E. coli crRNAs direct Cascade binding to >100 chromosomal sites. In contrast to the low specificity of Cascade-DNA interactions, >18 bp are required for both interference and primed adaptation. Hence, Cascade binding to suboptimal, off-target sites is inert. Our data support a model in which the initial Cascade association with DNA targets requires only limited sequence complementarity at the crRNA 5' end whereas recruitment and/or activation of the Cas3 nuclease, a prerequisite for interference and primed adaptation, requires extensive base pairing.IMPORTANCE Many bacterial and archaeal species encode CRISPR-Cas immunity systems that protect against invasion by foreign DNA. In the Escherichia coli CRISPR-Cas system, a protein complex, Cascade, binds 61-nucleotide (nt) CRISPR RNAs (crRNAs). The Cascade complex is directed to invading DNA molecules through base pairing between the crRNA and target DNA. This leads to recruitment of the Cas3 nuclease, which destroys the invading DNA molecule and promotes acquisition of new immunity elements. We made the first in vivo measurements of Cascade binding to DNA targets. Thus, we show that Cascade binding to DNA is highly promiscuous; endogenous E. coli crRNAs can direct Cascade binding to >100 chromosomal locations. In contrast, we show that targeted degradation and acquisition of new immunity elements require highly specific association of Cascade with DNA, limiting CRISPR-Cas function to the appropriate targets.
Keywords: CRISPR; Cas3; Cascade; PAM; priming; protospacer.
Copyright © 2018 Cooper et al.
Figures
References
-
- Makarova KS, Grishin NV, Shabalina SA, Wolf YI, Koonin EV. 2006. A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action. Biol Direct 1:7. doi: 10.1186/1745-6150-1-7. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
