

Androgen action in prostate function and disease
- PMID: 29666834
- PMCID: PMC5902724
Androgen action in prostate function and disease
Abstract
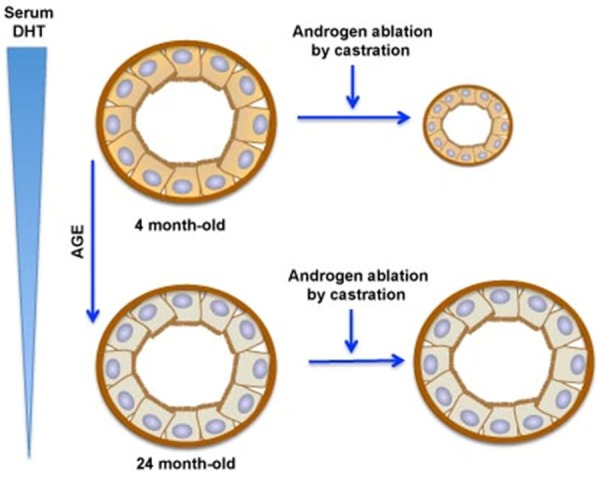
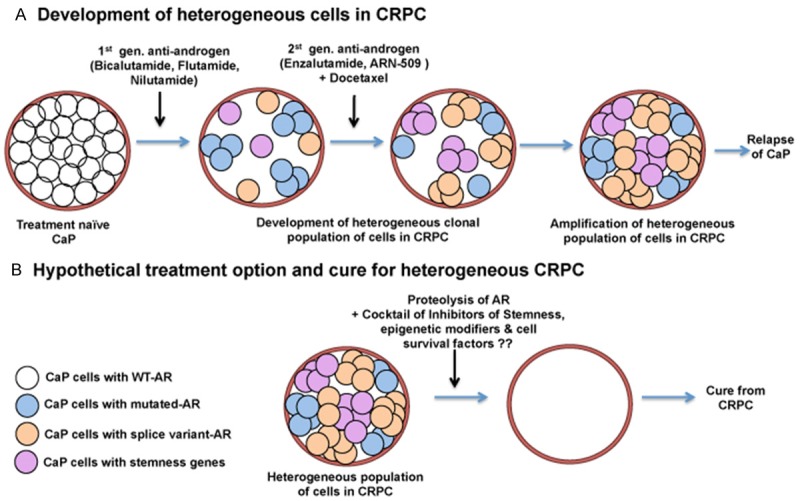
Benign prostatic hyperplasia (BPH) is an enlargement of the prostate gland that is frequently found in aging men. Androgens are essential for the development and differentiated function of the prostate, as well as for proliferation and survival of prostatic cells. In man, dog and rodent, there are age-related decreases in serum testosterone. Despite the lower serum testosterone levels, benign prostatic hyperplasia increases with age in men and dogs, while age-dependent prostatic hyperplasia develops in the dorsal and lateral lobes of the rat prostate. The possible mechanisms that lead to prostate hyperplasia have been extensively studied over many years. It is clear that androgens, estrogens and growth factors contribute to the condition, but the exact etiology remains unknown. Prostate cancer (CaP) represents a significant cause of death among males worldwide. As is the case of BPH, it is clear that androgens (testosterone and dihydrotestosterone) and their metabolites play important roles in the disease, but cause-effect relationships have not been established. Androgen deprivation therapy has been used for decades, primarily in the metastatic stage, to inhibit androgen-dependent prostate cancer cell growth. Androgen deprivation, which can be achieved by targeting hormone biosynthesis or androgen receptor activation, results in symptom amelioration. However, most patients will develop hormone refractory cancer or castration-resistant prostate cancer (CRPC). Prostatic epithelial cells demonstrate enormous plasticity in response to androgen ablation. This characteristic of prostatic epithelial cells may give rise to different populations of cells, some of which may not be dependent on androgen. Consequently, androgen receptor positive and negative cells might co-exist within CRPC. A clear understanding of this possible cellular heterogeneity and plasticity of prostate epithelial cells is necessary to develop an optimal strategy to treat or prevent CRPC.
Keywords: BPH; CRPC; Leydig cell; Prostate; aging; androgens.
Figures
References
-
- Habert R, Lejeune H, Saez JM. Origin, differentiation and regulation of fetal and adult Leydig cells. Mol Cell Endocrinol. 2001;179:47–74. - PubMed
-
- O’Shaughnessy PJ, Baker P, Sohnius U, Haavisto AM, Charlton HM, Huhtaniemi I. Fetal development of Leydig cell activity in the mouse is independent of pituitary gonadotroph function. Endocrinology. 1988;139:1141–1146. - PubMed
-
- Migrenne S, Pairault C, Racine C, Livera G, Géloso A, Habert R. Luteinizing hormone-dependent activity and luteinizing hormone-independent differentiation of rat fetal Leydig cells. Mol Cell Endocrinol. 2001;172:193–202. - PubMed
-
- Habert R, Picon R. Control of testicular steroidogenesis in foetal rat: effect of decapitation on testosterone and plasma luteinizing hormone-like activity. Acta Endocrinol. 1982;99:466–473. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous