

Diversity of Neuropeptide Cell-Cell Signaling Molecules Generated by Proteolytic Processing Revealed by Neuropeptidomics Mass Spectrometry
- PMID: 29667161
- PMCID: PMC5946320
- DOI: 10.1007/s13361-018-1914-1
Diversity of Neuropeptide Cell-Cell Signaling Molecules Generated by Proteolytic Processing Revealed by Neuropeptidomics Mass Spectrometry
Abstract
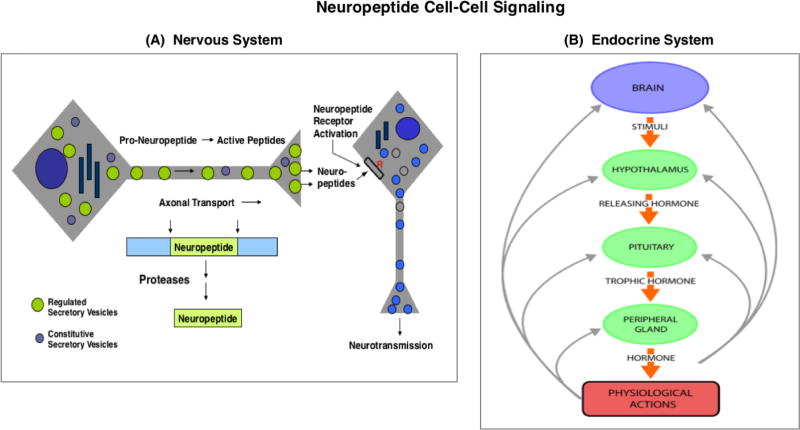
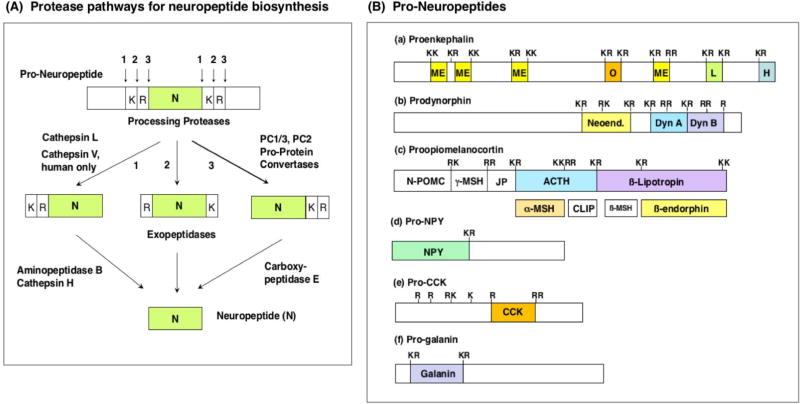
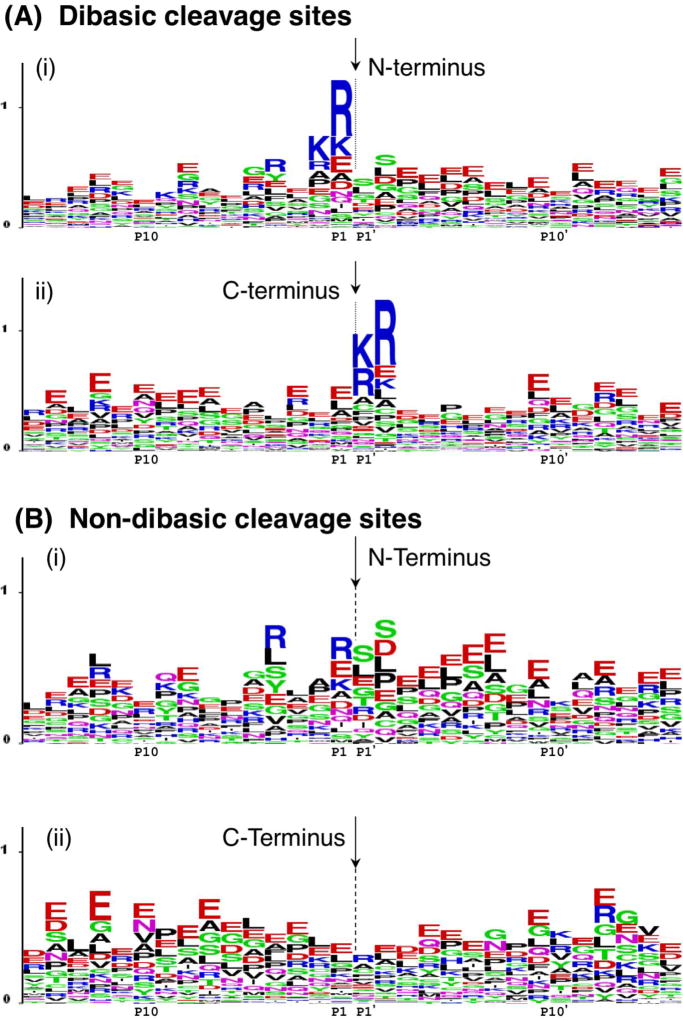
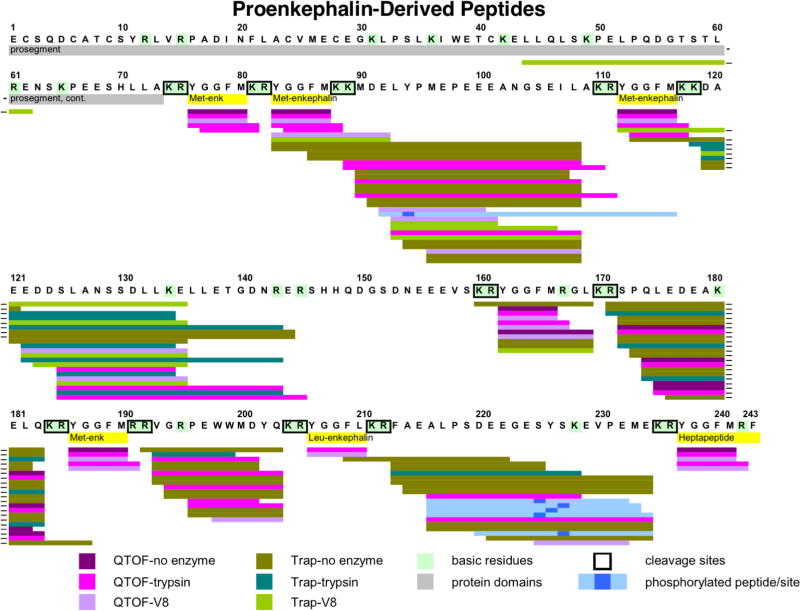
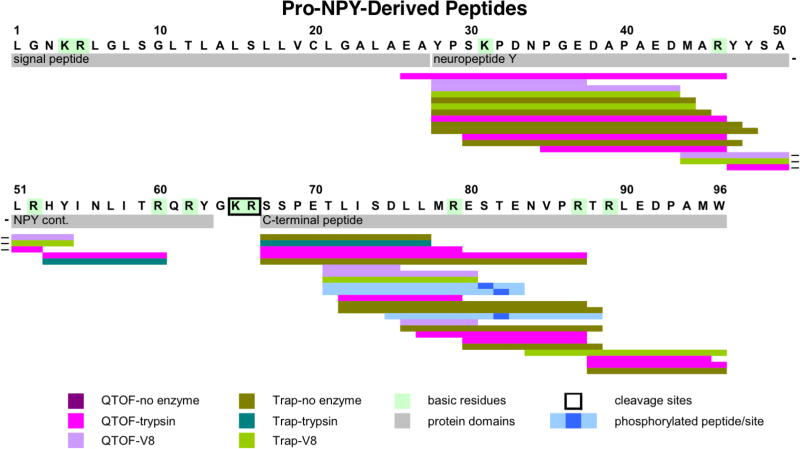
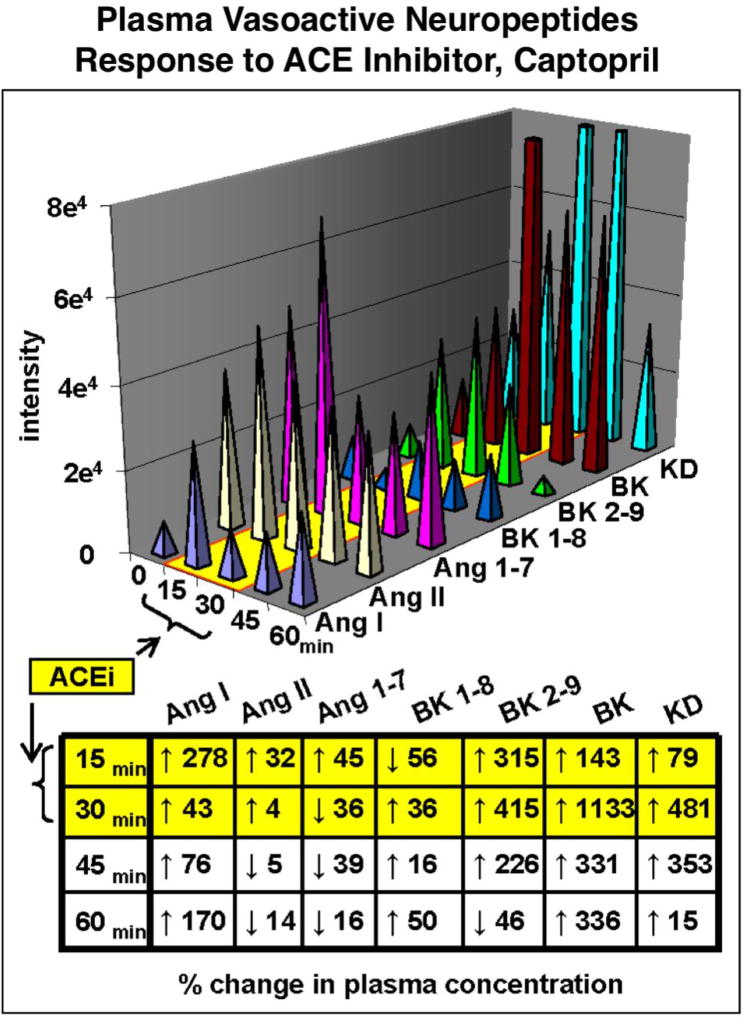
Neuropeptides are short peptides in the range of 3-40 residues that are secreted for cell-cell communication in neuroendocrine systems. In the nervous system, neuropeptides comprise the largest group of neurotransmitters. In the endocrine system, neuropeptides function as peptide hormones to coordinate intercellular signaling among target physiological systems. The diversity of neuropeptide functions is defined by their distinct primary sequences, peptide lengths, proteolytic processing of pro-neuropeptide precursors, and covalent modifications. Global, untargeted neuropeptidomics mass spectrometry is advantageous for defining the structural features of the thousands to tens of thousands of neuropeptides present in biological systems. Defining neuropeptide structures is the basis for defining the proteolytic processing pathways that convert pro-neuropeptides into active peptides. Neuropeptidomics has revealed that processing of pro-neuropeptides occurs at paired basic residues sites, and at non-basic residue sites. Processing results in neuropeptides with known functions and generates novel peptides representing intervening peptide domains flanked by dibasic residue processing sites, identified by neuropeptidomics. While very short peptide products of 2-4 residues are predicted from pro-neuropeptide dibasic processing sites, such peptides have not been readily identified; therefore, it will be logical to utilize metabolomics to identify very short peptides with neuropeptidomics in future studies. Proteolytic processing is accompanied by covalent post-translational modifications (PTMs) of neuropeptides comprising C-terminal amidation, N-terminal pyroglutamate, disulfide bonds, phosphorylation, sulfation, acetylation, glycosylation, and others. Neuropeptidomics can define PTM features of neuropeptides. In summary, neuropeptidomics for untargeted, global analyses of neuropeptides is essential for elucidation of proteases that generate diverse neuropeptides for cell-cell signaling. Graphical Abstract ᅟ.
Keywords: Cathepsin; Endocrine; Enkephalin; Mass spectrometry; NPY; Nervous system; Neuropeptides; Neurotransmission; Peptidomics; Pro-protein convertase; Proteases; Regulation.
Figures
References
-
- Kastin AJ. Handbook of biologically active peptides. Amsterdam: Elsevier; 2006.
-
- Siegel GJ, Albers RW, Fisher SK, Uhler MD. Basic Neurochemistry. Lippincott Williams and Wilkins; Philadelphia: 1999. pp. 363–382.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
