

Molecular Epidemiology of Extraintestinal Pathogenic Escherichia coli
- PMID: 29667573
- PMCID: PMC11575673
- DOI: 10.1128/ecosalplus.ESP-0004-2017
Molecular Epidemiology of Extraintestinal Pathogenic Escherichia coli
Abstract
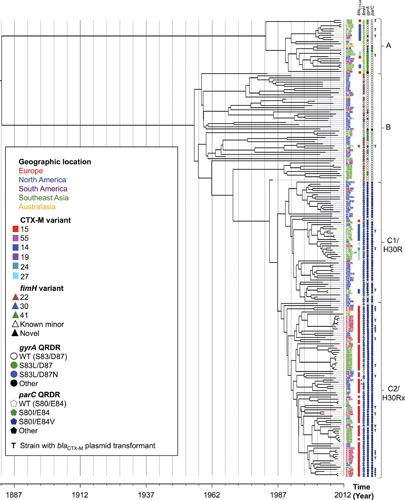
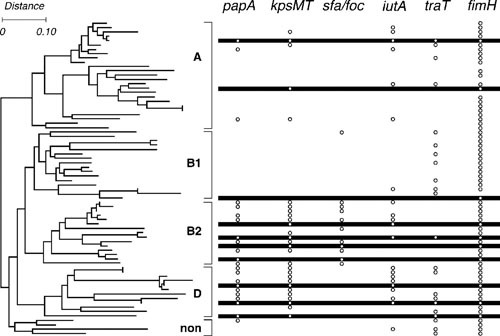
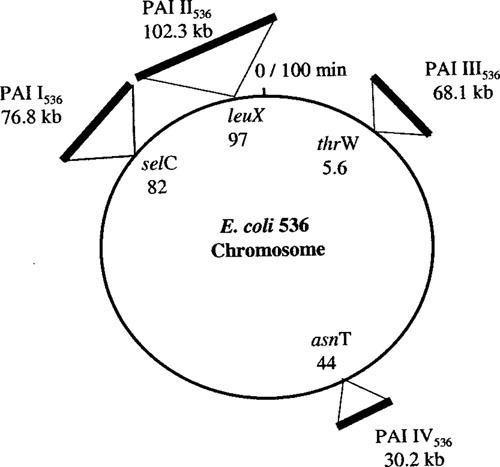
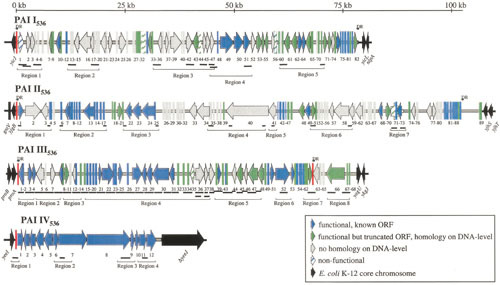
Extraintestinal pathogenic Escherichia coli (ExPEC) are important pathogens in humans and certain animals. Molecular epidemiological analyses of ExPEC are based on structured observations of E. coli strains as they occur in the wild. By assessing real-world phenomena as they occur in authentic contexts and hosts, they provide an important complement to experimental assessment. Fundamental to the success of molecular epidemiological studies are the careful selection of subjects and the use of appropriate typing methods and statistical analysis. To date, molecular epidemiological studies have yielded numerous important insights into putative virulence factors, host-pathogen relationships, phylogenetic background, reservoirs, antimicrobial-resistant strains, clinical diagnostics, and transmission pathways of ExPEC, and have delineated areas in which further study is needed. The rapid pace of discovery of new putative virulence factors and the increasing awareness of the importance of virulence factor regulation, expression, and molecular variation should stimulate many future molecular epidemiological investigations. The growing sophistication and availability of molecular typing methodologies, and of the new computational and statistical approaches that are being developed to address the huge amounts of data that whole genome sequencing generates, provide improved tools for such studies and allow new questions to be addressed.
Figures
References
-
- Russo TA, Johnson JR. 2003. Medical and economic impact of extraintestinal infections due to Escherichia coli: an overlooked epidemic. Microbes Infect 5:449–456. - PubMed
-
- Lipsky BA. 1989. Urinary tract infections in men. Epidemiology, pathophysiology, diagnosis, and treatment. Ann Intern Med 110:138–150. - PubMed
-
- Andriole VT, Patterson TF. 1991. Epidemiology, natural history, and management of urinary tract infections in pregnancy. Med Clin North Am 75:359–373. - PubMed
-
- Rothman KJ, Greenland S. 1998. The Emergence of Modern Epidemiology, 2nd ed. Lippincott-Raven, Philadelphia.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
