

DNA-mediated dimerization on a compact sequence signature controls enhancer engagement and regulation by FOXA1
- PMID: 29669022
- PMCID: PMC6009666
- DOI: 10.1093/nar/gky259
DNA-mediated dimerization on a compact sequence signature controls enhancer engagement and regulation by FOXA1
Abstract
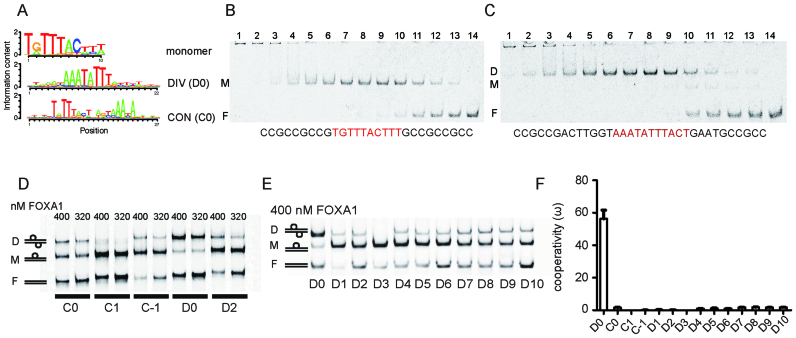
FOXA1 is a transcription factor capable to bind silenced chromatin to direct context-dependent cell fate conversion. Here, we demonstrate that a compact palindromic DNA element (termed 'DIV' for its diverging half-sites) induces the homodimerization of FOXA1 with strongly positive cooperativity. Alternative structural models are consistent with either an indirect DNA-mediated cooperativity or a direct protein-protein interaction. The cooperative homodimer formation is strictly constrained by precise half-site spacing. Re-analysis of chromatin immunoprecipitation sequencing data indicates that the DIV is effectively targeted by FOXA1 in the context of chromatin. Reporter assays show that FOXA1-dependent transcriptional activity declines when homodimeric binding is disrupted. In response to phosphatidylinositol-3 kinase inhibition DIV sites pre-bound by FOXA1 such as at the PVT1/MYC locus exhibit a strong increase in accessibility suggesting a role of the DIV configuration in the chromatin closed-open dynamics. Moreover, several disease-associated single nucleotide polymorphisms map to DIV elements and show allelic differences in FOXA1 homodimerization, reporter gene expression and are annotated as quantitative trait loci. This includes the rs541455835 variant at the MAPT locus encoding the Tau protein associated with Parkinson's disease. Collectively, the DIV guides chromatin engagement and regulation by FOXA1 and its perturbation could be linked to disease etiologies.
Figures





References
-
- Cirillo L.A., Lin F.R., Cuesta I., Friedman D., Jarnik M., Zaret K.S.. Opening of compacted chromatin by early developmental transcription factors HNF3 (FoxA) and GATA-4. Mol. Cell. 2002; 9:279–289. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
