

An Antibody Targeting the Fusion Machinery Neutralizes Dual-Tropic Infection and Defines a Site of Vulnerability on Epstein-Barr Virus
- PMID: 29669253
- PMCID: PMC5909843
- DOI: 10.1016/j.immuni.2018.03.026
An Antibody Targeting the Fusion Machinery Neutralizes Dual-Tropic Infection and Defines a Site of Vulnerability on Epstein-Barr Virus
Abstract
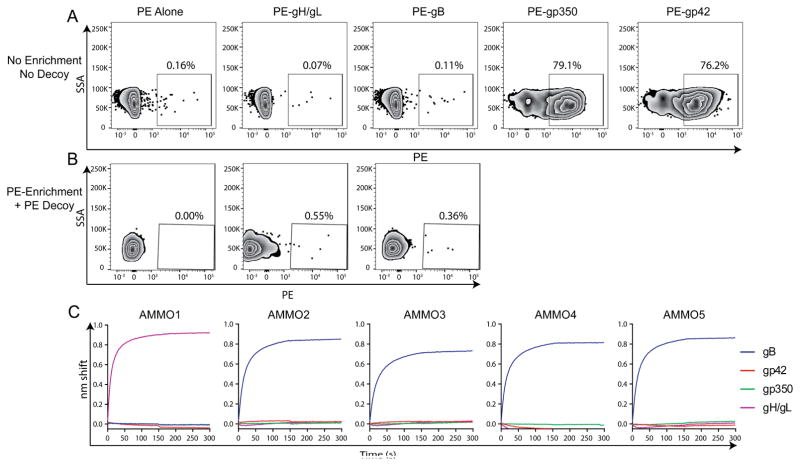
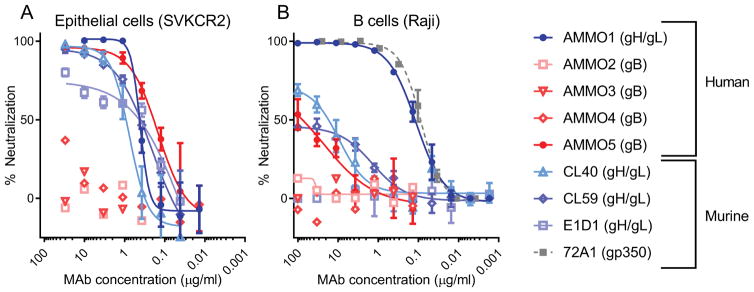
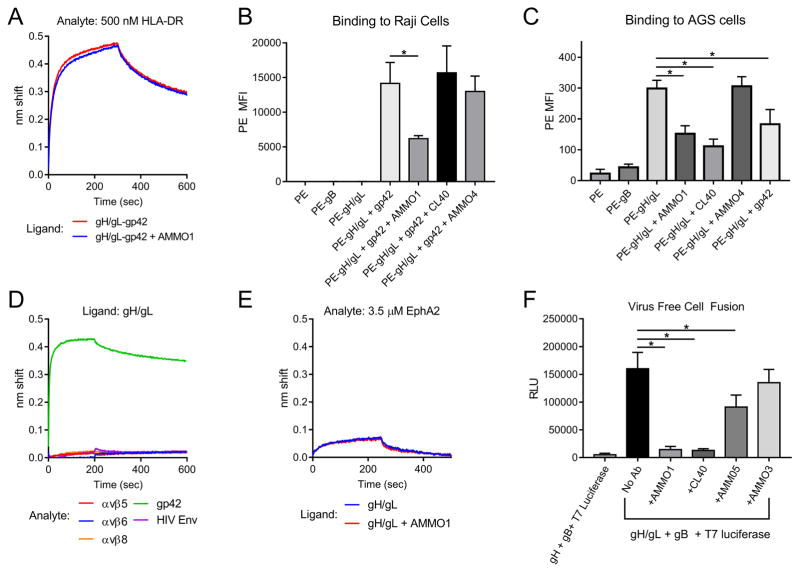
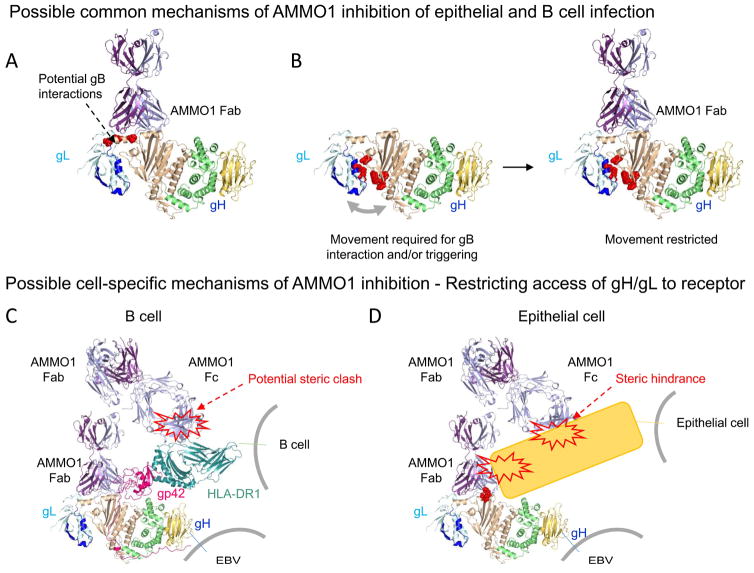
Epstein-Barr virus (EBV) is a causative agent of infectious mononucleosis and is associated with 200,000 new cases of cancer and 140,000 deaths annually. Subunit vaccines against this pathogen have focused on the gp350 glycoprotein and remain unsuccessful. We isolated human antibodies recognizing the EBV fusion machinery (gH/gL and gB) from rare memory B cells. One anti-gH/gL antibody, AMMO1, potently neutralized infection of B cells and epithelial cells, the two major cell types targeted by EBV. We determined a cryo-electron microscopy reconstruction of the gH/gL-gp42-AMMO1 complex and demonstrated that AMMO1 bound to a discontinuous epitope formed by both gH and gL at the Domain-I/Domain-II interface. Integrating structural, biochemical, and infectivity data, we propose that AMMO1 inhibits fusion of the viral and cellular membranes. This work identifies a crucial epitope that may aid in the design of next-generation subunit vaccines against this major public health burden.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
A.T.M discloses that he has filed provisional patents, 62/504,447 and 62/560,061 related to this work.
Figures



Comment in
-
Inhibiting EBV: The Key to Carefree Smooching.Immunity. 2018 Apr 17;48(4):623-625. doi: 10.1016/j.immuni.2018.03.036. Immunity. 2018. PMID: 29669245
-
To EBV or not to EBV: Rational vaccine design for a common infection.Sci Immunol. 2018 May 4;3(23):eaat9661. doi: 10.1126/sciimmunol.aat9661. Sci Immunol. 2018. PMID: 29728427
References
-
- Agirre J, Iglesias-Fernandez J, Rovira C, Davies GJ, Wilson KS, Cowtan KD. Privateer: software for the conformational validation of carbohydrate structures. Nature structural & molecular biology. 2015;22:833–834. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
