

TMEM150C/Tentonin3 Is a Regulator of Mechano-gated Ion Channels
- PMID: 29669276
- PMCID: PMC5929159
- DOI: 10.1016/j.celrep.2018.03.094
TMEM150C/Tentonin3 Is a Regulator of Mechano-gated Ion Channels
Abstract
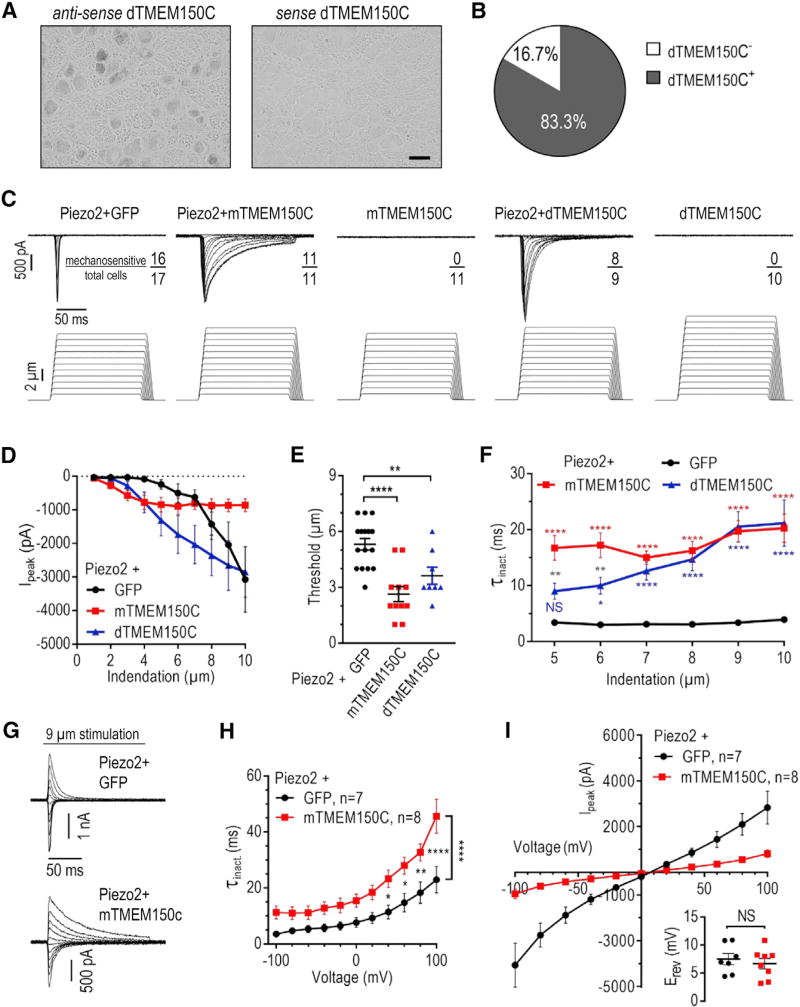
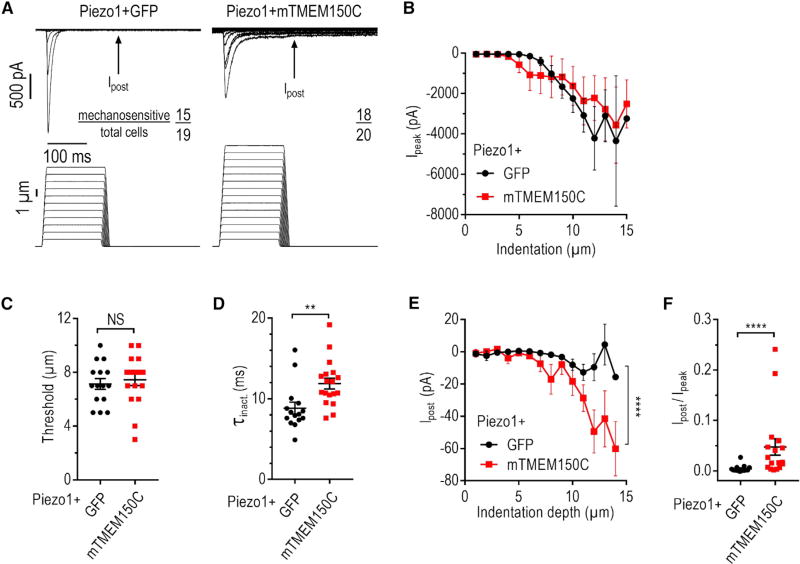
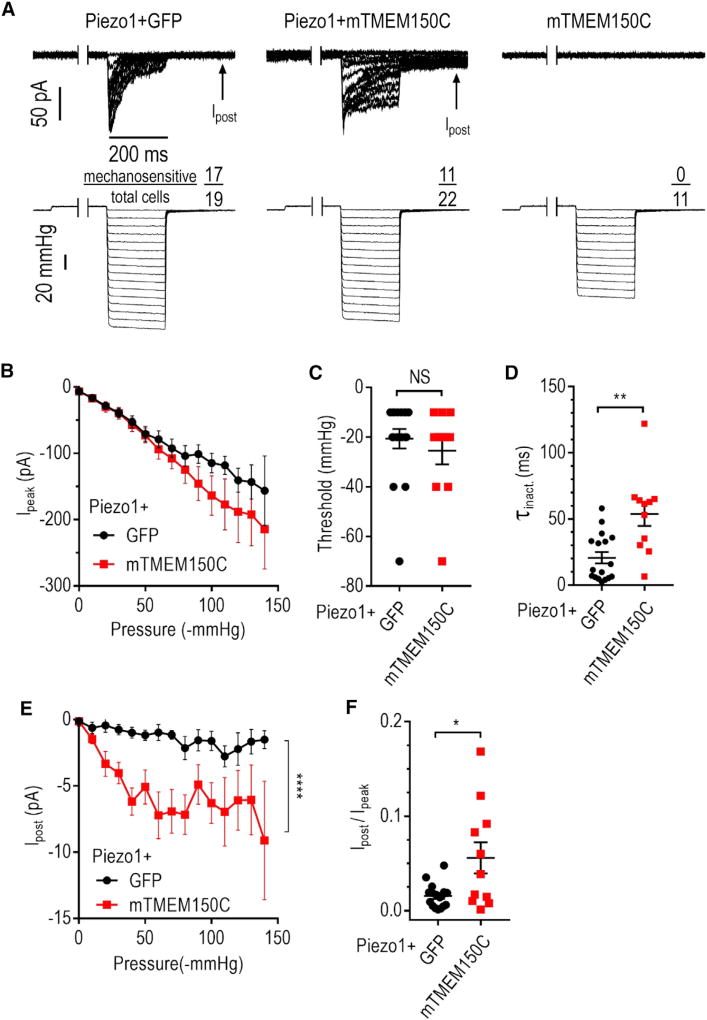
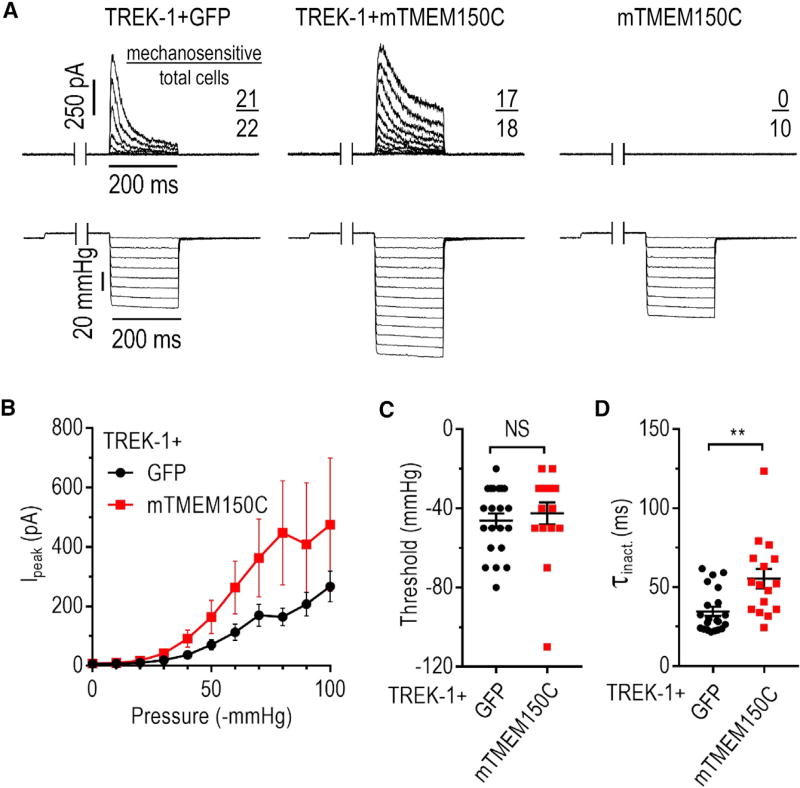
Neuronal mechano-sensitivity relies on mechano-gated ion channels, but pathways regulating their activity remain poorly understood. TMEM150C was proposed to mediate mechano-activated current in proprioceptive neurons. Here, we studied functional interaction of TMEM150C with mechano-gated ion channels from different classes (Piezo2, Piezo1, and the potassium channel TREK-1) using two independent methods of mechanical stimulation. We found that TMEM150C significantly prolongs the duration of the mechano-current produced by all three channels, decreases apparent activation threshold in Piezo2, and induces persistent current in Piezo1. We also show that TMEM150C is co-expressed with Piezo2 in trigeminal neurons, expanding its role beyond proprioceptors. Finally, we cloned TMEM150C from the trigeminal neurons of the tactile-foraging domestic duck and showed that it functions similarly to the mouse ortholog, demonstrating evolutionary conservation among vertebrates. Our studies reveal TMEM150C as a general regulator of mechano-gated ion channels from different classes.
Keywords: K2P2.1; Piezo1; Piezo2; TMEM150C; TREK-1; Tentonin3; mechano-receptor; mechano-sensitivity; mechano-transduction; trigeminal ganglia.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Albuisson J, Murthy SE, Bandell M, Coste B, Louis-Dit-Picard H, Mathur J, Fénéant-Thibault M, Tertian G, de Jaureguiberry JP, Syfuss PY, et al. Dehydrated hereditary stomatocytosis linked to gain-of-function mutations in mechanically activated PIEZO1 ion channels. Nat. Commun. 2013;4:1884. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
